

mRNA 5'-leader trans-splicing in the chordates
- PMID: 11159910
- PMCID: PMC312621
- DOI: 10.1101/gad.865401
mRNA 5'-leader trans-splicing in the chordates
Abstract
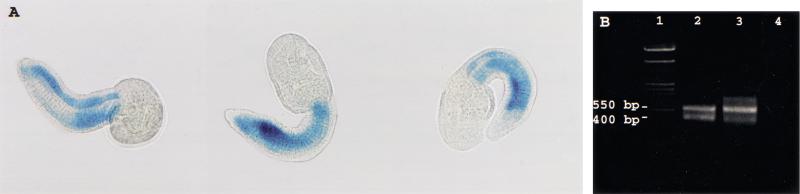
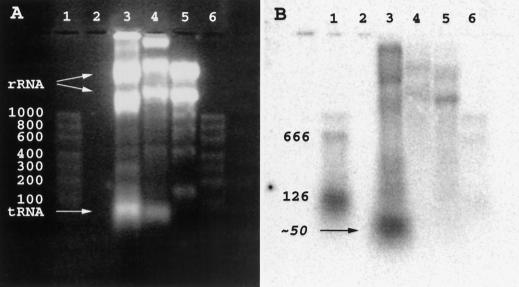
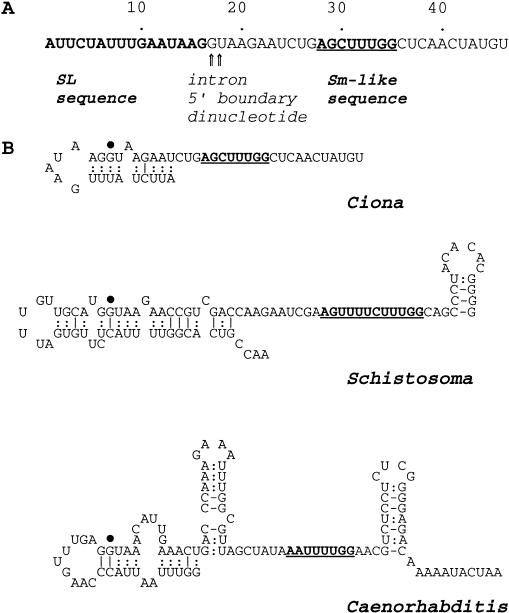
We report the discovery of mRNA 5'-leader trans-splicing (SL trans-splicing) in the chordates. In the ascidian protochordate Ciona intestinalis, the mRNAs of at least seven genes undergo trans-splicing of a 16-nucleotide 5'-leader apparently derived from a 46-nucleotide RNA that shares features with previously characterized splice donor SL RNAs. SL trans-splicing was known previously to occur in several protist and metazoan phyla, however, this is the first report of SL trans-splicing within the deuterostome division of the metazoa. SL trans-splicing is not known to occur in the vertebrates. However, because ascidians are primitive chordates related to vertebrate ancestors, our findings raise the possibility of ancestral SL trans-splicing in the vertebrate lineage.
Figures




References
-
- Agabian N. Trans splicing of nuclear pre-mRNAs. Cell. 1990;61:1157–1160. - PubMed
-
- Akopian AN, Okuse K, Souslova V, England S, Ogata N, Wood JN. Trans-splicing of a voltage-gated sodium channel is regulated by nerve growth factor. FEBS Lett. 1999;445:177–182. - PubMed
-
- Ausoni S, Campione M, Picard A, Moretti P, Vitadello M, De Nardi C, Schiaffino S. Structure and regulation of the mouse cardiac troponin I gene. J Biol Chem. 1994;269:339–346. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources