

Glucocorticoids upregulate CD40 ligand expression and induce CD40L-dependent immunoglobulin isotype switching
- PMID: 11160161
- PMCID: PMC199190
- DOI: 10.1172/JCI10168
Glucocorticoids upregulate CD40 ligand expression and induce CD40L-dependent immunoglobulin isotype switching
Abstract
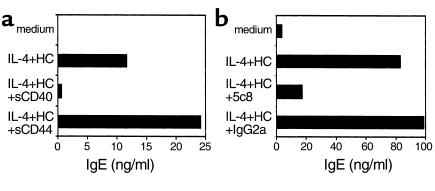
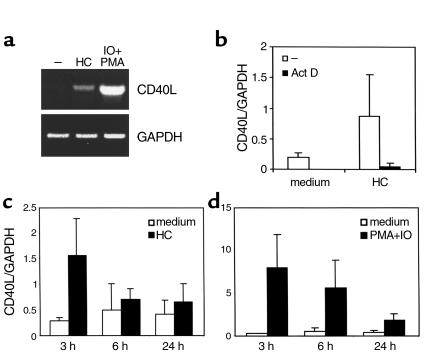
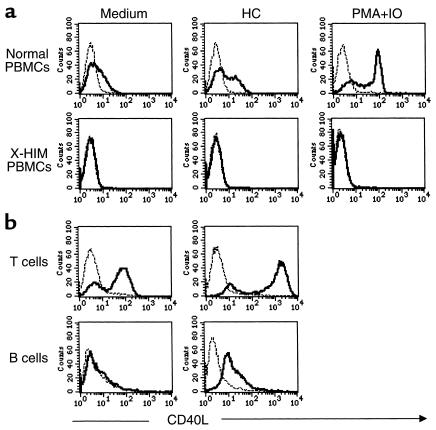
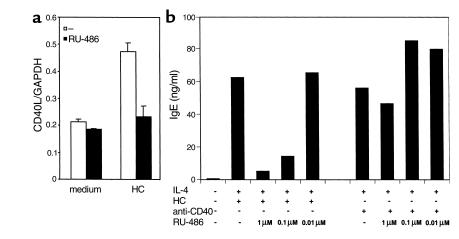
IL-4 and CD40 ligation are essential for IgE synthesis by B cells. We have shown previously that hydrocortisone (HC) induces IgE synthesis in IL-4-stimulated human B cells. In this study we demonstrate that HC fails to induce IgE synthesis in B cells from CD40 ligand-deficient (CD40L-deficient) patients. Disruption of CD40L-CD40 interactions by soluble CD40-Ig fusion protein or anti-CD40L mAb blocked the capacity of HC to induce IgE synthesis in normal B cells. HC upregulated CD40L mRNA expression in PBMCs and surface expression of CD40L in PBMCs as well as in purified populations of T and B cells. Upregulation of CD40L mRNA in PBMCs occurred 3 hours after stimulation with HC and was inhibited by actinomycin D. Upregulation of CD40L mRNA and induction of IgE synthesis by HC were inhibited by the steroid hormone receptor antagonist RU-486. These results indicate that ligand-mediated activation of the glucocorticoid receptor upregulates CD40L expression in human lymphocytes.
Figures
Comment on
-
Corticosteroids, IgE, and atopy.J Clin Invest. 2001 Feb;107(3):265-6. doi: 10.1172/JCI12157. J Clin Invest. 2001. PMID: 11160147 Free PMC article. No abstract available.
References
-
- Vercelli D, Geha RS. Regulation of isotype switching. Curr Opin Immunol. 1992; 4:794–799. - PubMed
-
- Zurawski S, et al. The primary binding subunit of the human interleukin-4 receptor is also a component of the interleukin-13 receptor. J Biol Chem. 1995; 270:13869–13878. - PubMed
-
- Vercelli D, Geha RS. Regulation of IgE synthesis: from the membrane to the genes. Springer Semin Immunopathol. 1993; 15:5–16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
