

Late oligodendrocyte progenitors coincide with the developmental window of vulnerability for human perinatal white matter injury
- PMID: 11160401
- PMCID: PMC6762224
- DOI: 10.1523/JNEUROSCI.21-04-01302.2001
Late oligodendrocyte progenitors coincide with the developmental window of vulnerability for human perinatal white matter injury
Abstract
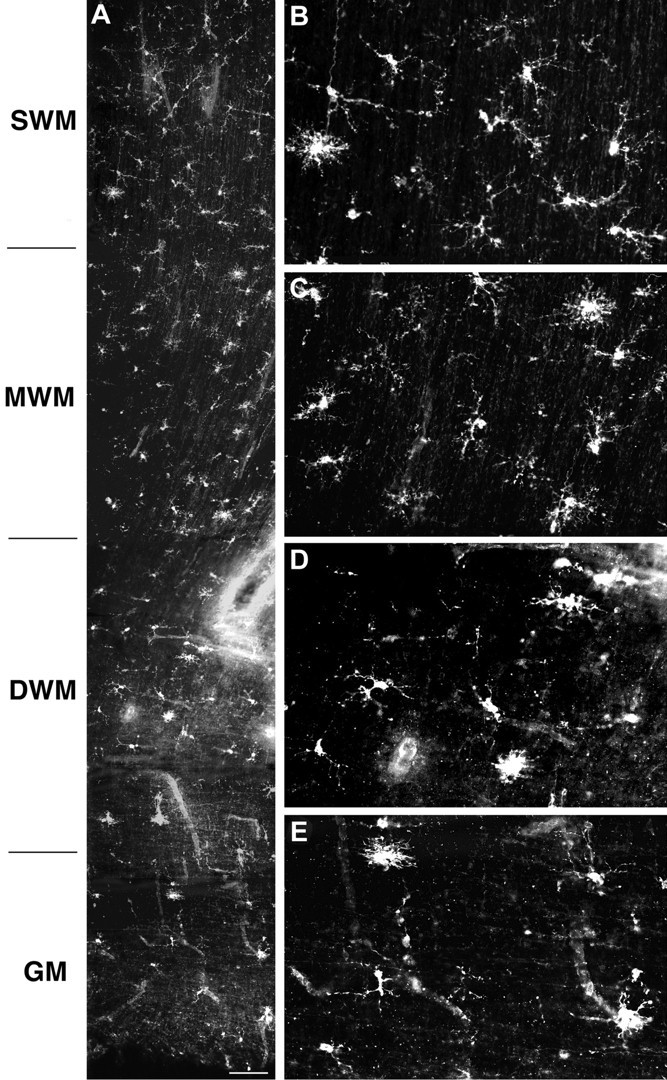
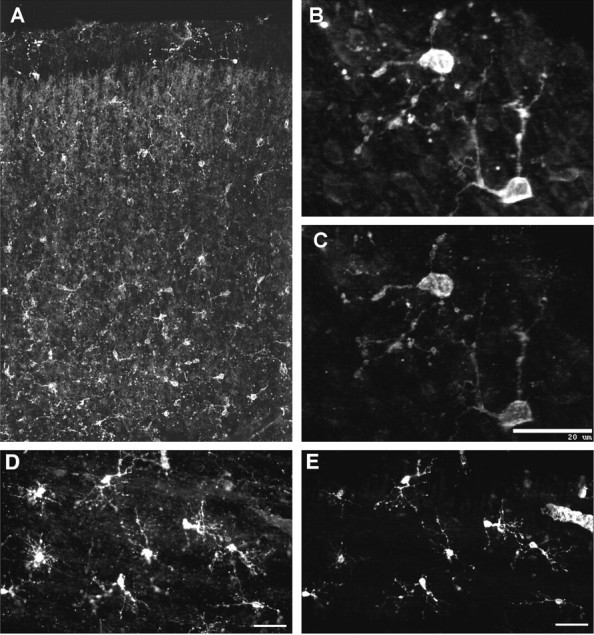
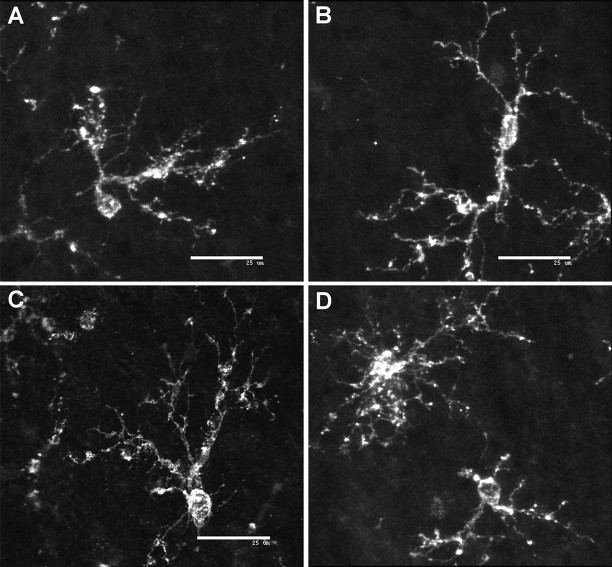
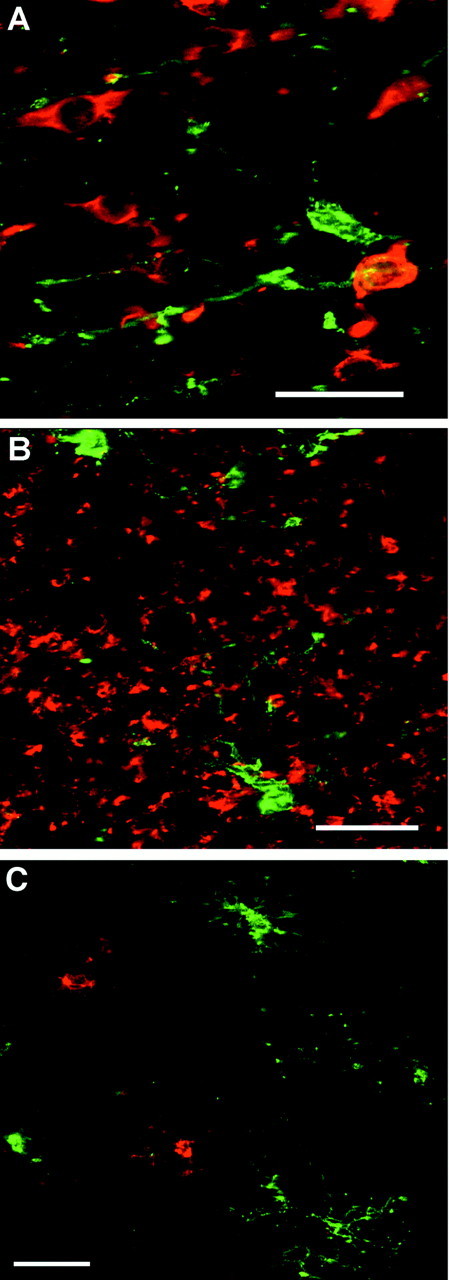
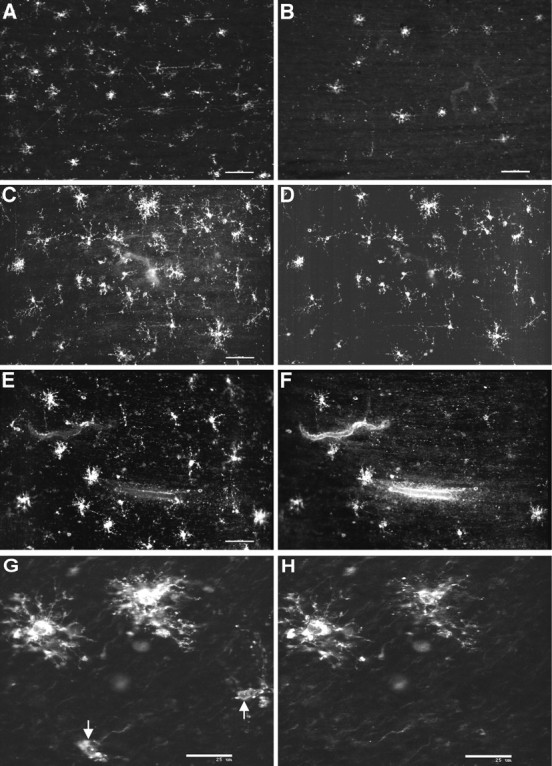
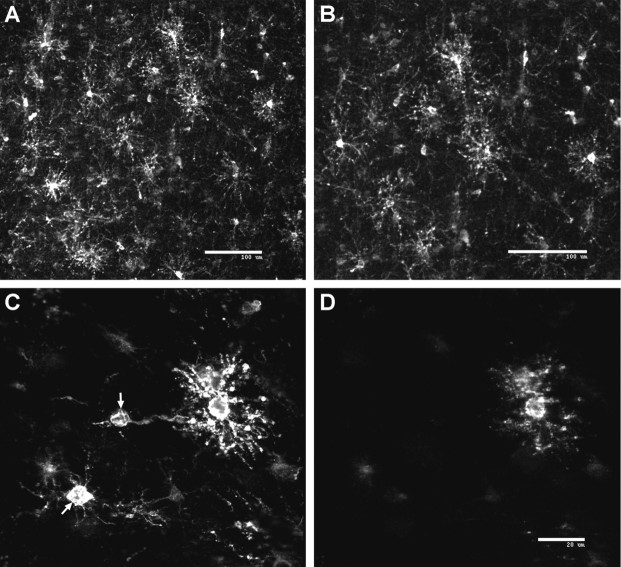
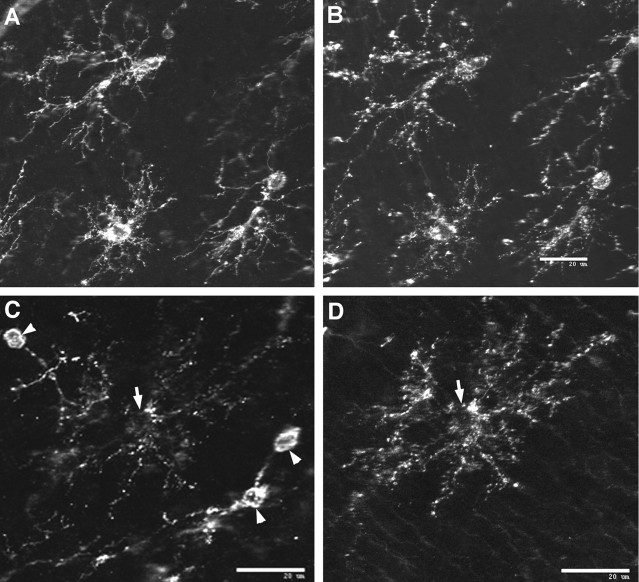
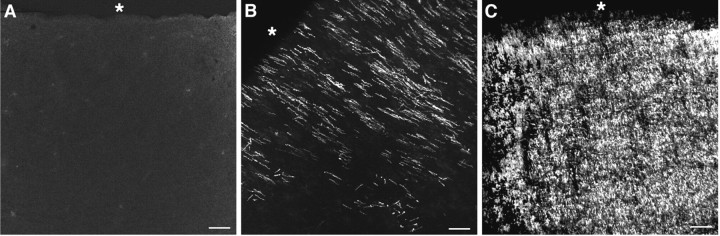
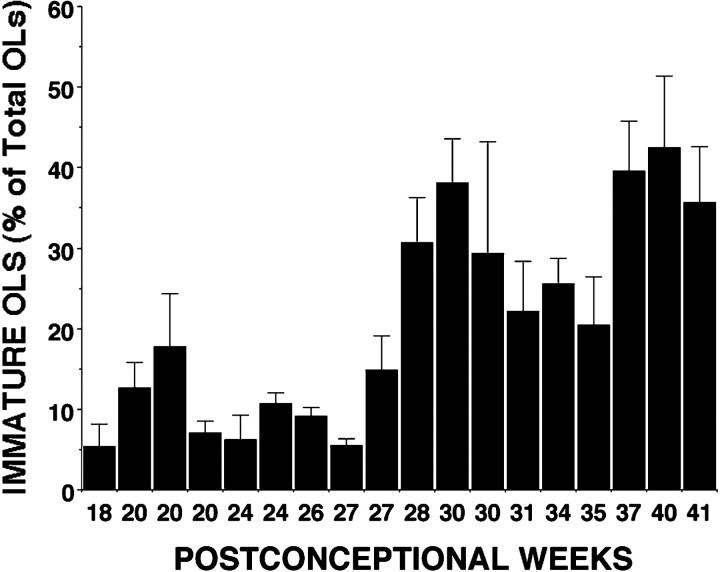
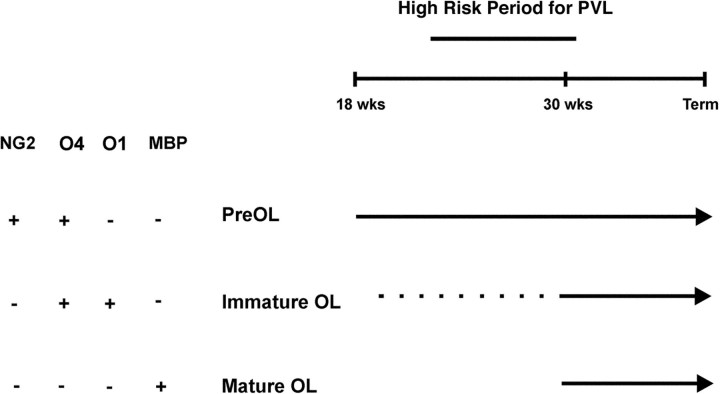
Hypoxic-ischemic injury to the periventricular cerebral white matter [periventricular leukomalacia (PVL)] results in cerebral palsy and is the leading cause of brain injury in premature infants. The principal feature of PVL is a chronic disturbance of myelination and suggests that oligodendrocyte (OL) lineage progression is disrupted by ischemic injury. We determined the OL lineage stages at risk for injury during the developmental window of vulnerability for PVL (23-32 weeks, postconceptional age). In 26 normal control autopsy human brains, OL lineage progression was defined in parietal white matter, a region of predilection for PVL. Three successive OL stages, the late OL progenitor, the immature OL, and the mature OL, were characterized between 18 and 41 weeks with anti-NG2 proteoglycan, O4, O1, and anti-myelin basic protein (anti-MBP) antibodies. NG2+O4+ late OL progenitors were the predominant stage throughout the latter half of gestation. Between 18 and 27 weeks, O4+O1+ immature OLs were a minor population (9.9 +/- 2.1% of total OLs; n = 9). Between 28 and 41 weeks, an increase in immature OLs to 30.9 +/- 2.1% of total OLs (n = 9) was accompanied by a progressive increase in MBP+ myelin sheaths that were restricted to the periventricular white matter. The developmental window of high risk for PVL thus precedes the onset of myelination and identifies the late OL progenitor as the major potential target. Moreover, the decline in incidence of PVL at approximately 32 weeks coincides with the onset of myelination in the periventricular white matter and suggests that the risk for PVL is related to the presence of late OL progenitors in the periventricular white matter.
Figures
References
-
- Back SA, Volpe JJ. Cellular and molecular pathogenesis of periventricular white matter injury. Ment Retard Dev Disabil Res Rev. 1997;3:96–107.
-
- Back SA, Volpe JJ. Approaches to the study of diseases involving oligodendroglial death. In: Koliatsos V, Ratan RR, editors. Cell death and diseases of the nervous system. Humana; Totowa, NJ: 1999. pp. 401–428.
-
- Bansal R, Warrington AE, Gard A, Ranscht B, Pfeiffer SE. Multiple and novel specificities of monoclonal antibodies O1, O4, and R-mAb used in the analysis of oligodendrocyte development. J Neurosci Res. 1989;24:548–557. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous