

Reliability of macaque frontal eye field neurons signaling saccade targets during visual search
- PMID: 11160450
- PMCID: PMC6763809
- DOI: 10.1523/JNEUROSCI.21-02-00713.2001
Reliability of macaque frontal eye field neurons signaling saccade targets during visual search
Abstract
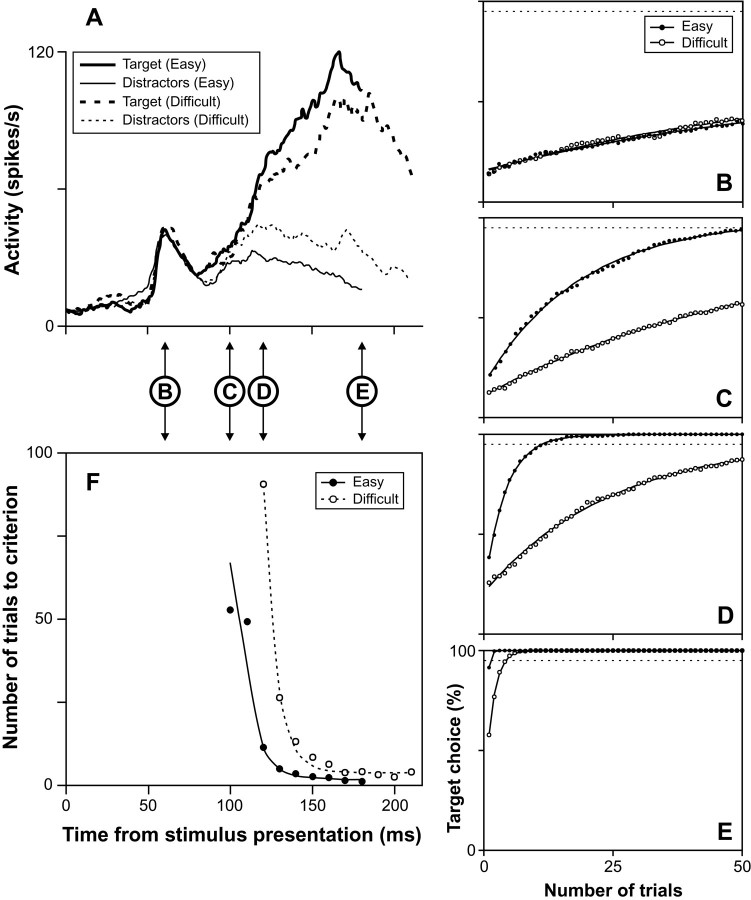
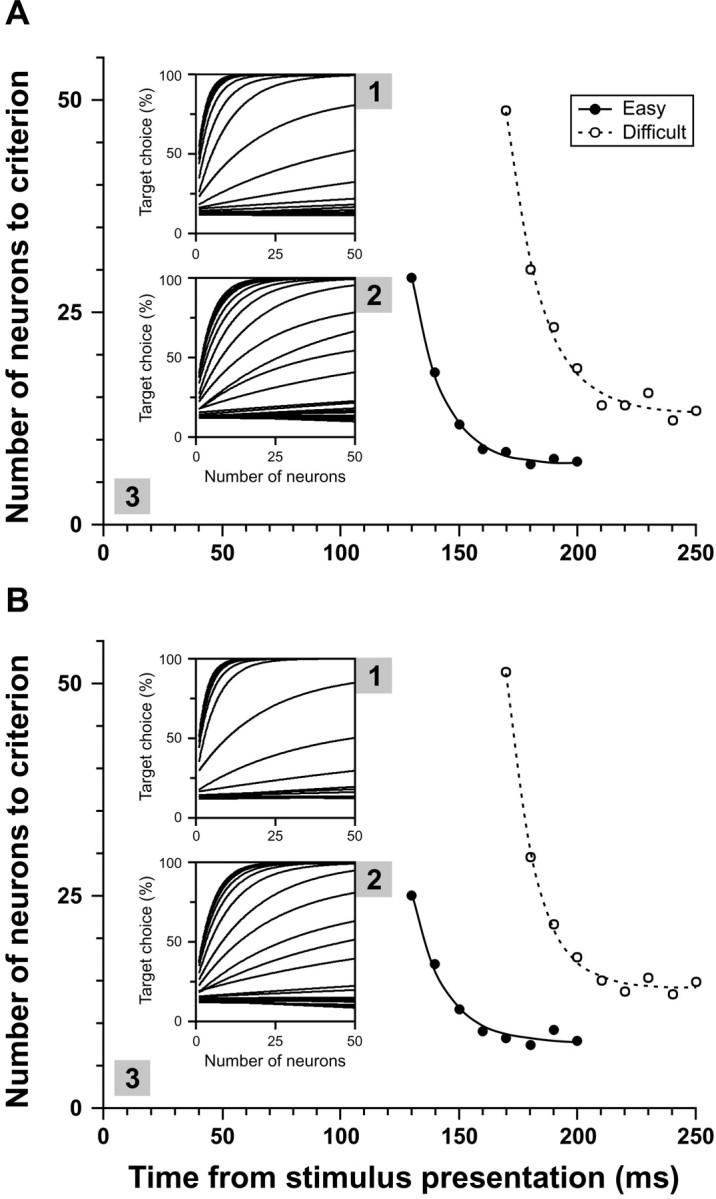
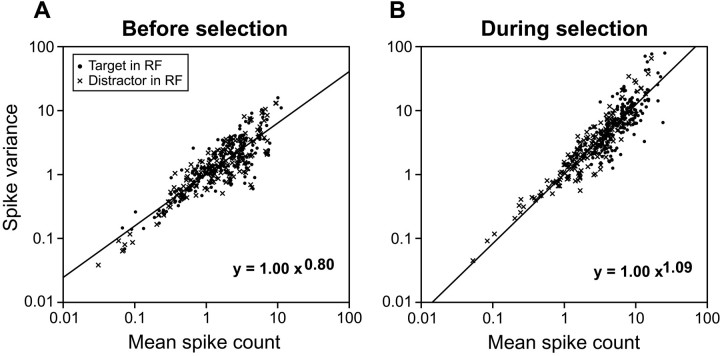
Although many studies have explored the neural correlates of visual attention and selection, few have examined the reliability with which neurons represent relevant information. We monitored activity in the frontal eye field (FEF) of monkeys trained to make a saccade to a target defined by the conjunction of color and shape or to a target defined by color differences. The difficulty of conjunction search was manipulated by varying the number of distractors, and the difficulty of feature search was manipulated by varying the similarity in color between target and distractors. The reliability of individual neurons in signaling the target location in correct trials was determined using a neuron-anti-neuron approach within a winner-take-all architecture. On average, approximately seven trials of the activity of single neurons were sufficient to match near-perfect behavioral performance in the easiest search, and approximately 14 trials were sufficient in the most difficult search. We also determined how many neurons recorded separately need to be evaluated within a trial to match behavioral performance. Results were quantitatively similar to those of the single neuron analysis. We also found that signal reliability in the FEF did not change with task demands, and overall, behavioral accuracy across the search tasks was approximated when only six trials or neurons were combined. Furthermore, whether combining trials or neurons, the increase in time of target discrimination corresponded to the increase in mean saccade latency across visual search difficulty levels. Finally, the variance of spike counts in the FEF increased as a function of the mean spike count, and the parameters of this relationship did not change with attentional selection.
Figures







References
-
- Bichot NP. Attention, eye movements, and neurons: linking physiology and behavior. In: Harris LR, Jenkin MRM, editors. Vision and attention. Springer-Verlag; New York: 2001.
-
- Bichot NP, Schall JD. Saccade target selection in macaque during feature and conjunction visual search. Vis Neurosci. 1999a;16:81–89. - PubMed
-
- Bichot NP, Schall JD. Effects of similarity and history on neural mechanisms of visual selection. Nat Neurosci. 1999b;2:549–554. - PubMed
-
- Bichot NP, Schall JD, Thompson KG. Visual feature selectivity in frontal eye fields induced by experience in mature macaques. Nature. 1996;381:697–699. - PubMed
-
- Britten KH, Shadlen MN, Newsome WT, Movshon JA. Responses of neurons in macaque MT to stochastic motion signals. Vis Neurosci. 1993;10:1157–1169. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources