

Serine phosphorylation of focal adhesion kinase in interphase and mitosis: a possible role in modulating binding to p130(Cas)
- PMID: 11160818
- PMCID: PMC30563
- DOI: 10.1091/mbc.12.1.1
Serine phosphorylation of focal adhesion kinase in interphase and mitosis: a possible role in modulating binding to p130(Cas)
Abstract
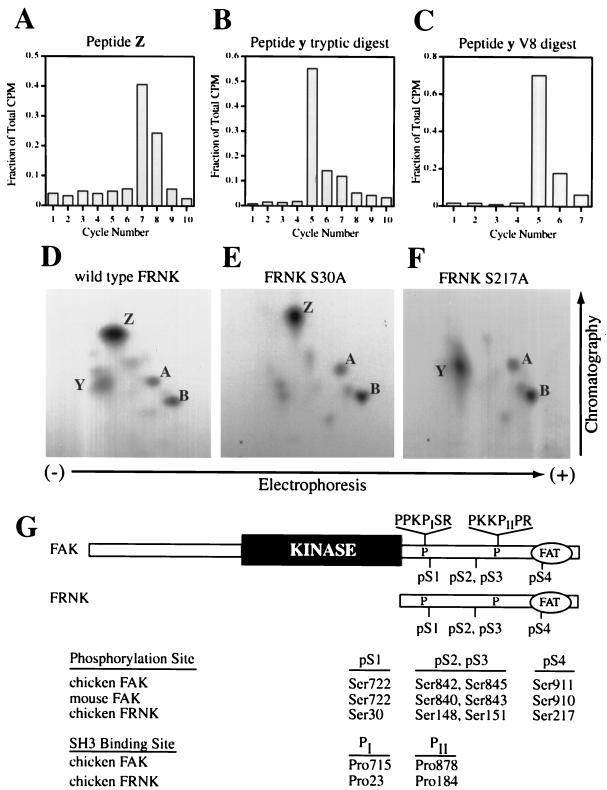
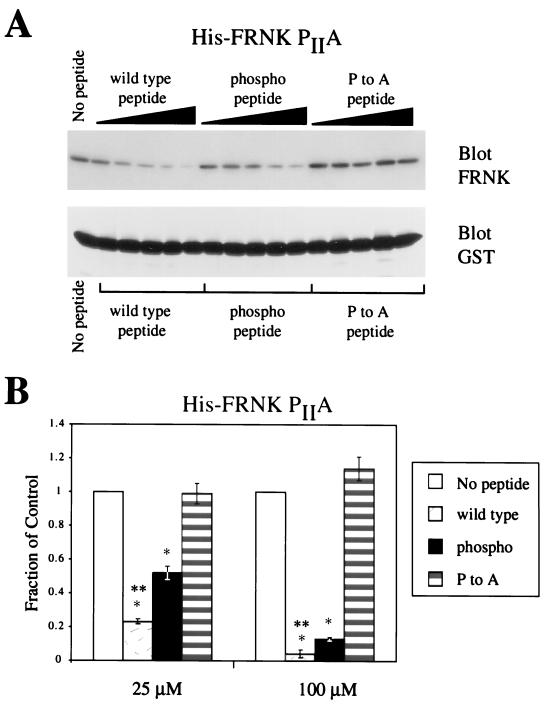
Focal adhesion kinase (FAK) is an important regulator of integrin signaling in adherent cells and accordingly its activity is significantly modulated during mitosis when cells detach from the extracellular matrix. During mitosis, FAK becomes heavily phosphorylated on serine residues concomitant with its inactivation and dephosphorylation on tyrosine. Little is known about the regulation of FAK activity by serine phosphorylation. In this report, we characterize two novel sites of serine phosphorylation within the C-terminal domain of FAK. Phosphorylation-specific antibodies directed to these sites and against two previously characterized sites of serine phosphorylation were used to study the regulated phosphorylation of FAK in unsynchronized and mitotic cells. Among the four major phosphorylation sites, designated pS1-pS4, phosphorylation of pS1 (Ser722) is unchanged in unsynchronized and mitotic cells. In contrast, pS3 and pS4 (Ser843 and Ser910) exhibit increased phosphorylation during mitosis. In vitro peptide binding experiments provide evidence that phosphorylation of pS1 (Ser722) may play a role in modulating FAK binding to the SH3 domain of the adapter protein p130(Cas).
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
