

Thermostable and site-specific DNA binding of the gene product ORF56 from the Sulfolobus islandicus plasmid pRN1, a putative archael plasmid copy control protein
- PMID: 11160922
- PMCID: PMC29613
- DOI: 10.1093/nar/29.4.904
Thermostable and site-specific DNA binding of the gene product ORF56 from the Sulfolobus islandicus plasmid pRN1, a putative archael plasmid copy control protein
Abstract
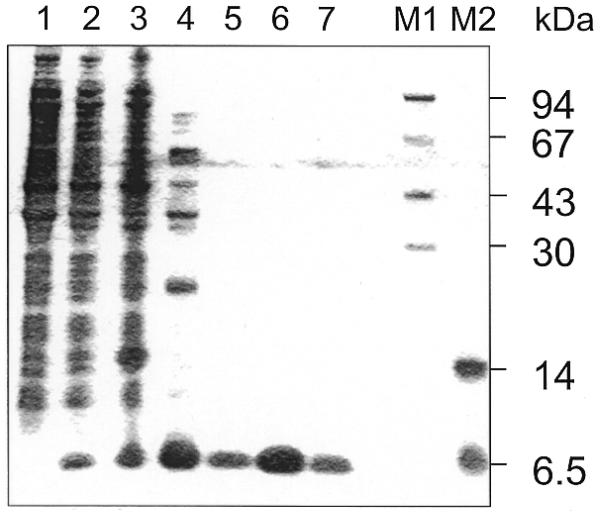
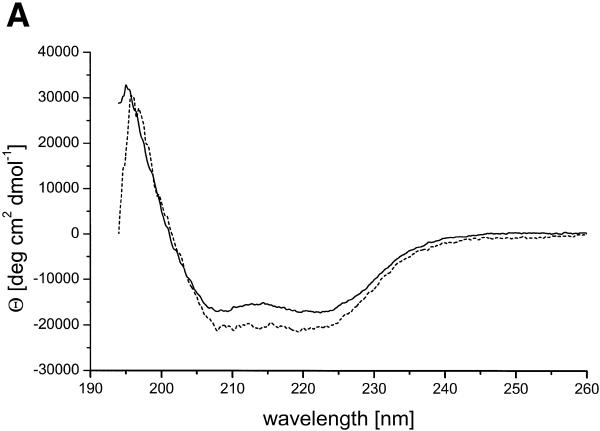
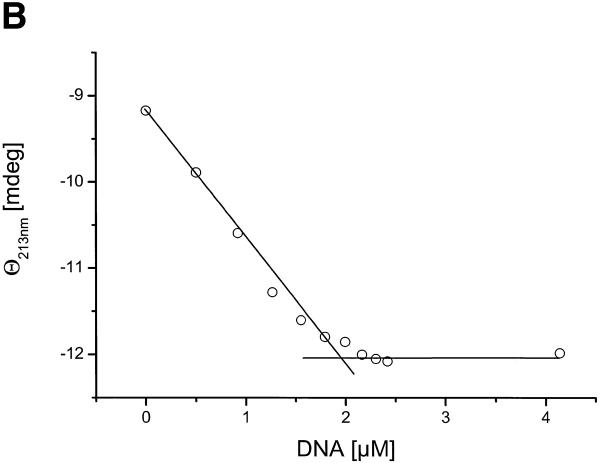
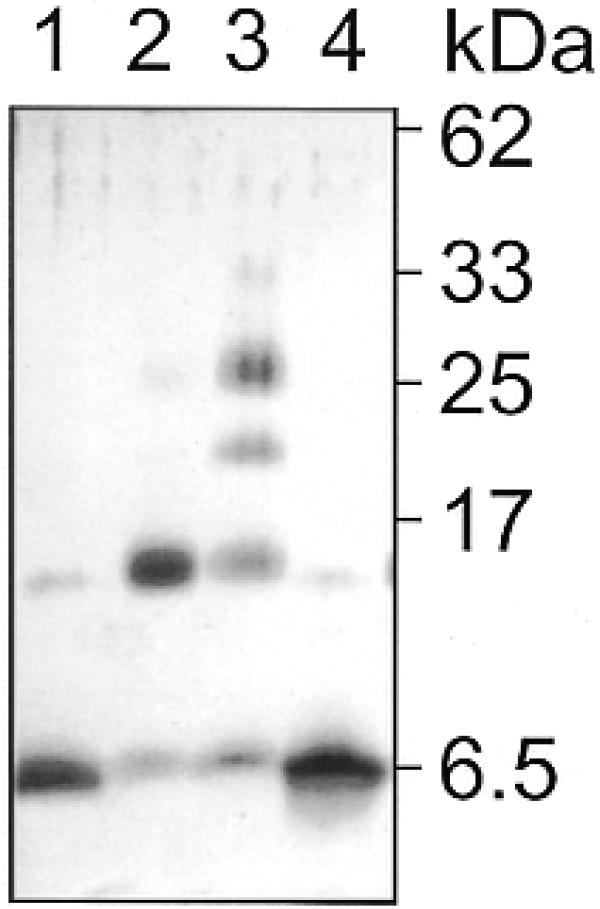
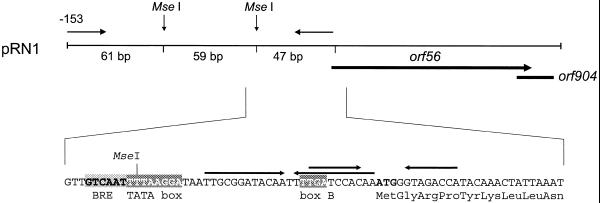
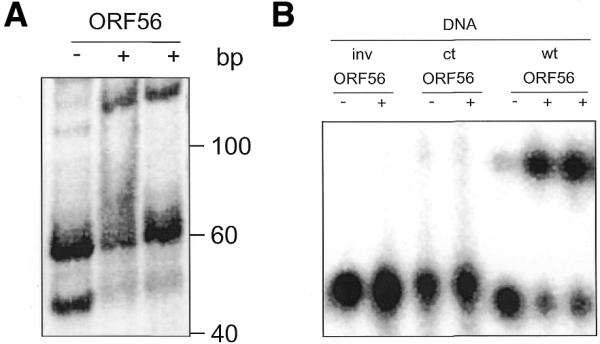
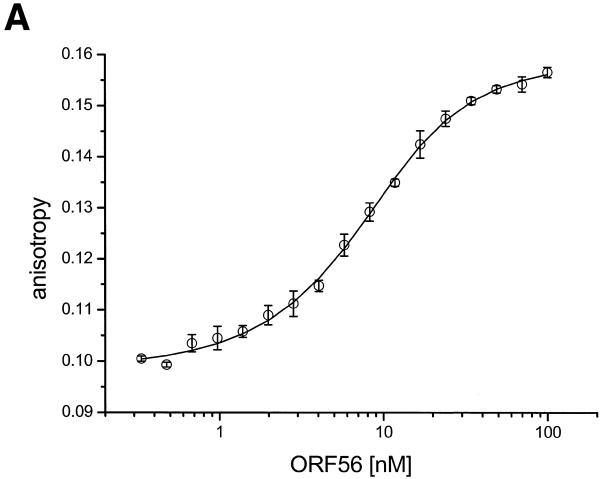
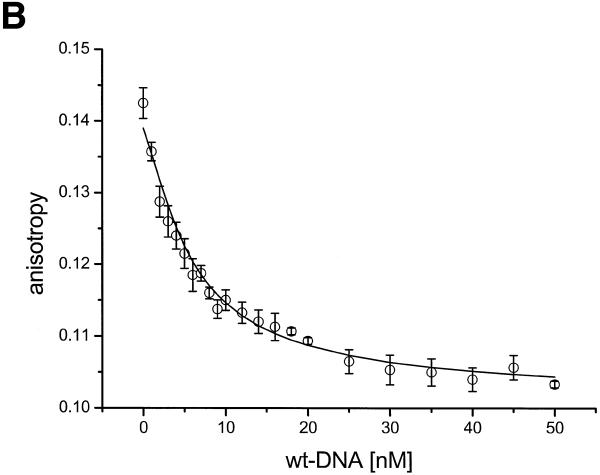
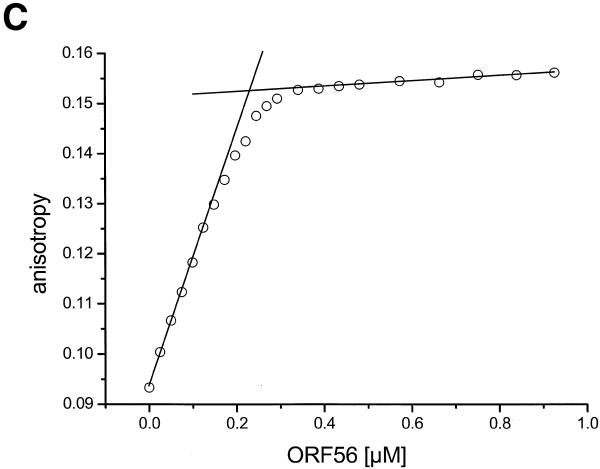
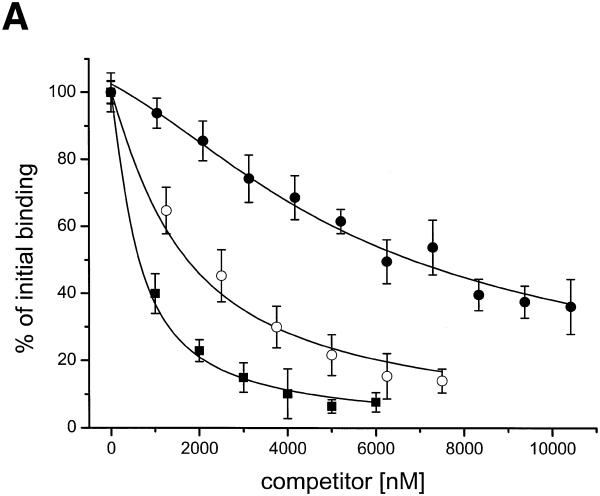
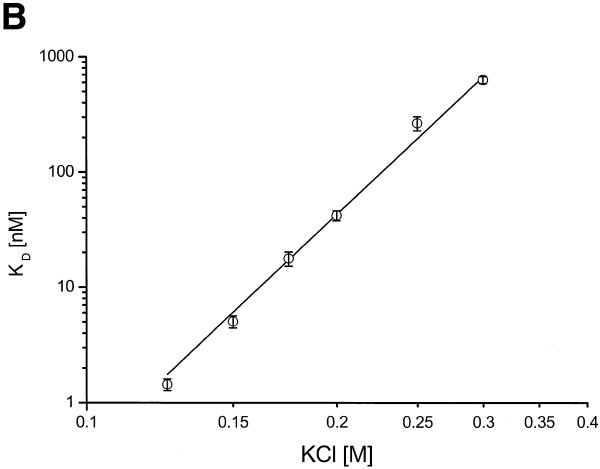
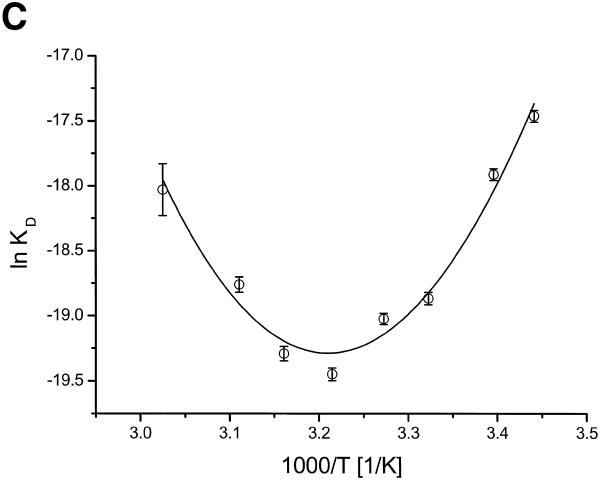
There is still a lack of information on the specific characteristics of DNA-binding proteins from hyperthermophiles. Here we report on the product of the gene orf56 from plasmid pRN1 of the acidophilic and thermophilic archaeon Sulfolobus islandicus. orf56 has not been characterised yet but low sequence similarily to several eubacterial plasmid-encoded genes suggests that this 6.5 kDa protein is a sequence-specific DNA-binding protein. The DNA-binding properties of ORF56, expressed in Escherichia coli, have been investigated by EMSA experiments and by fluorescence anisotropy measurements. Recombinant ORF56 binds to double-stranded DNA, specifically to an inverted repeat located within the promoter of orf56. Binding to this site could down-regulate transcription of the orf56 gene and also of the overlapping orf904 gene, encoding the putative initiator protein of plasmid replication. By gel filtration and chemical crosslinking we have shown that ORF56 is a dimeric protein. Stoichiometric fluorescence anisotropy titrations further indicate that ORF56 binds as a tetramer to the inverted repeat of its target binding site. CD spectroscopy points to a significant increase in ordered secondary structure of ORF56 upon binding DNA. ORF56 binds without apparent cooperativity to its target DNA with a dissociation constant in the nanomolar range. Quantitative analysis of binding isotherms performed at various salt concentrations and at different temperatures indicates that approximately seven ions are released upon complex formation and that complex formation is accompanied by a change in heat capacity of -6.2 kJ/mol. Furthermore, recombinant ORF56 proved to be highly thermostable and is able to bind DNA up to 85 degrees C.
Figures

Similar articles
-
The protein ORF80 from the acidophilic and thermophilic archaeon Sulfolobus islandicus binds highly site-specifically to double-stranded DNA and represents a novel type of basic leucine zipper protein.Nucleic Acids Res. 2001 Dec 15;29(24):4973-82. doi: 10.1093/nar/29.24.4973. Nucleic Acids Res. 2001. PMID: 11812827 Free PMC article.
-
Structure-based stability analysis of an extremely stable dimeric DNA binding protein from Sulfolobus islandicus.Biochemistry. 2009 Oct 27;48(42):10030-7. doi: 10.1021/bi900760n. Biochemistry. 2009. PMID: 19788170
-
Folding and association of an extremely stable dimeric protein from Sulfolobus islandicus.J Mol Biol. 2004 Feb 6;336(1):227-40. doi: 10.1016/j.jmb.2003.12.003. J Mol Biol. 2004. PMID: 14741218
-
The replication protein of the Sulfolobus islandicus plasmid pRN1.Biochem Soc Trans. 2004 Apr;32(Pt 2):240-4. doi: 10.1042/bst0320240. Biochem Soc Trans. 2004. PMID: 15046580 Review.
-
Signal peptides of secreted proteins of the archaeon Sulfolobus solfataricus: a genomic survey.Arch Microbiol. 2002 Mar;177(3):209-16. doi: 10.1007/s00203-001-0386-y. Epub 2002 Jan 11. Arch Microbiol. 2002. PMID: 11907676 Review.
Cited by
-
Temperature dependence and thermodynamics of Klenow polymerase binding to primed-template DNA.Biophys J. 2006 Mar 1;90(5):1739-51. doi: 10.1529/biophysj.105.071837. Epub 2005 Dec 9. Biophys J. 2006. PMID: 16339886 Free PMC article.
-
Transcriptional analysis of the genetic element pSSVx: differential and temporal regulation of gene expression reveals correlation between transcription and replication.J Bacteriol. 2007 Sep;189(17):6339-50. doi: 10.1128/JB.00638-07. Epub 2007 Jun 22. J Bacteriol. 2007. PMID: 17586636 Free PMC article.
-
Genomic comparison of archaeal conjugative plasmids from Sulfolobus.Archaea. 2004 Oct;1(4):231-9. doi: 10.1155/2004/151926. Archaea. 2004. PMID: 15810432 Free PMC article.
-
A genetically economical family of plasmid-encoded transcriptional repressors involved in control of plasmid copy number.J Bacteriol. 2002 Sep;184(18):4943-51. doi: 10.1128/JB.184.18.4943-4951.2002. J Bacteriol. 2002. PMID: 12193609 Free PMC article. Review. No abstract available.
-
A novel type of replicative enzyme harbouring ATPase, primase and DNA polymerase activity.EMBO J. 2003 May 15;22(10):2516-25. doi: 10.1093/emboj/cdg246. EMBO J. 2003. PMID: 12743045 Free PMC article.
References
-
- Zillig W., Kletzin,A., Schleper,C., Holz,I., Janekovic,D., Hain,J., Lanzendoerfer,M. and Kristjansson,J.K. (1994) Screening for Sulfolobales, their plasmids and their viruses in Icelandic solfataras. Syst. Appl. Microbiol., 16, 609–628.
-
- Keeling P.J., Klenk,H.P., Singh,R.K., Feeley,O., Schleper,C., Zillig,W., Doolittle,W.F. and Sensen,C.W. (1996) Complete nucleotide sequence of the Sulfolobus islandicus multicopy plasmid pRN1. Plasmid, 35, 141–144. - PubMed
-
- Peng X., Holz,I., Zillig,W., Garrett,R.A. and She,Q. (2000) Evolution of the family of pRN plasmids and their integrase-mediated insertion into the chromosome of the crenarchaeon Sulfolobus solfataricus. J. Mol. Biol., 303, 449–454. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
