

Protein-RNA interactions: a structural analysis
- PMID: 11160927
- PMCID: PMC29619
- DOI: 10.1093/nar/29.4.943
Protein-RNA interactions: a structural analysis
Abstract
A detailed computational analysis of 32 protein-RNA complexes is presented. A number of physical and chemical properties of the intermolecular interfaces are calculated and compared with those observed in protein-double-stranded DNA and protein-single-stranded DNA complexes. The interface properties of the protein-RNA complexes reveal the diverse nature of the binding sites. van der Waals contacts played a more prevalent role than hydrogen bond contacts, and preferential binding to guanine and uracil was observed. The positively charged residue, arginine, and the single aromatic residues, phenylalanine and tyrosine, all played key roles in the RNA binding sites. A comparison between protein-RNA and protein-DNA complexes showed that whilst base and backbone contacts (both hydrogen bonding and van der Waals) were observed with equal frequency in the protein-RNA complexes, backbone contacts were more dominant in the protein-DNA complexes. Although similar modes of secondary structure interactions have been observed in RNA and DNA binding proteins, the current analysis emphasises the differences that exist between the two types of nucleic acid binding protein at the atomic contact level.
Figures
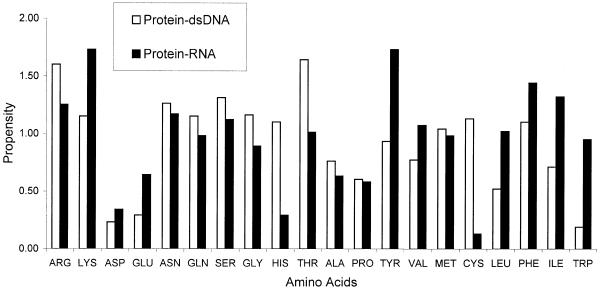
References
-
- Moore P.B. (1998) The three-dimensional structure of the ribosome and its components. Annu. Rev. Biophys. Biomol. Struct., 27, 35–58. - PubMed
-
- Ramakrishnan V. and White,S.W. (1998) Ribosomal protein structures: insights into the architecture, machinery and evolution of the ribosome. Trends Biochem. Sci., 23, 208–212. - PubMed
-
- Luhrmann R., Kastner,B. and Bach,M. (1990) Structure of spliceosomal snRNP’s and their role in pre-mRNA splicing. Biochim. Biophys. Acta, 1087, 265–292. - PubMed
-
- Tarasow T.M. and Eaton,B.E. (1998) Dressed for success: realising the catalytic potential of RNA. Biopolymers ,48, 29–37.
-
- Scott W.G. and Klug,A. (1996) Ribozymes: structures and mechanism in RNA catalysis. Trends Biochem. Sci., 21, 220–224. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
