

Vascular endothelial growth factor-C-mediated lymphangiogenesis promotes tumour metastasis
- PMID: 11179212
- PMCID: PMC145430
- DOI: 10.1093/emboj/20.4.672
Vascular endothelial growth factor-C-mediated lymphangiogenesis promotes tumour metastasis
Abstract
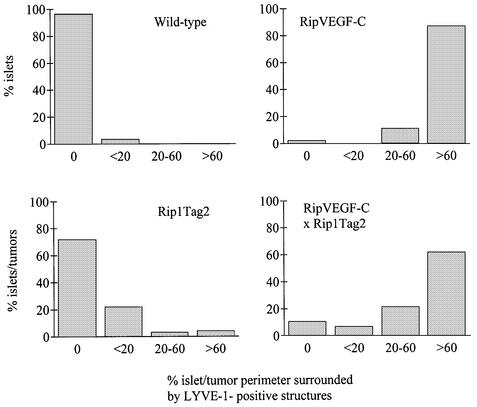
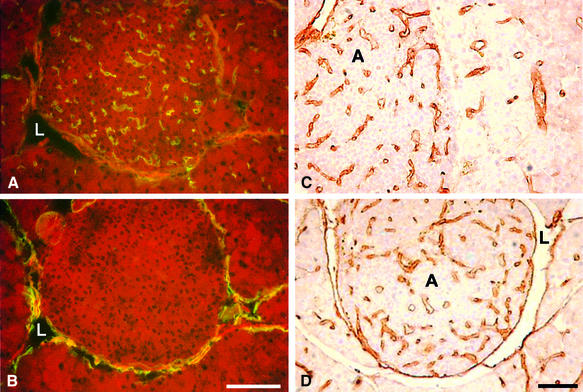
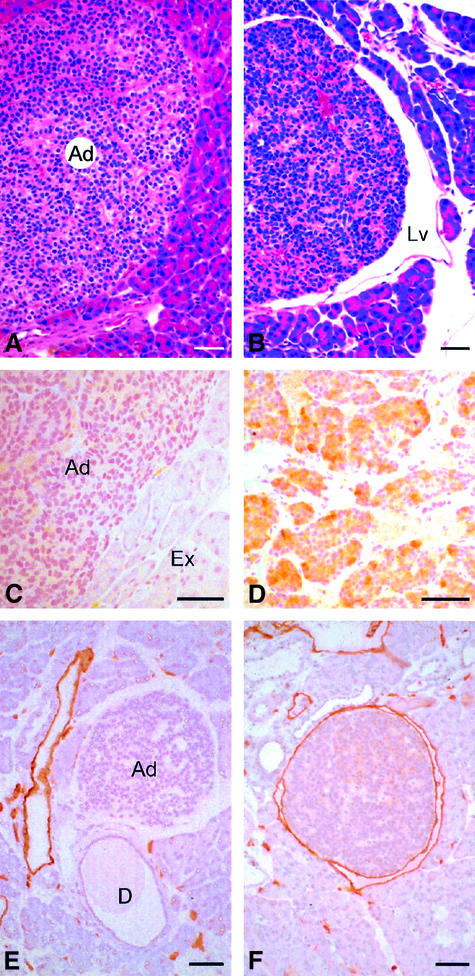
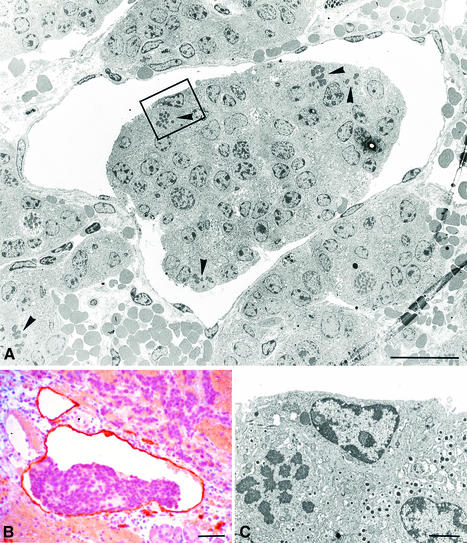
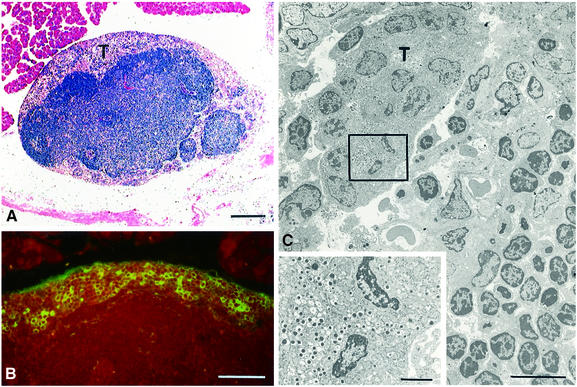
Metastasis is a frequent and lethal complication of cancer. Vascular endothelial growth factor-C (VEGF-C) is a recently described lymphangiogenic factor. Increased expression of VEGF-C in primary tumours correlates with dissemination of tumour cells to regional lymph nodes. However, a direct role for VEGF-C in tumour lymphangiogenesis and subsequent metastasis has yet to be demonstrated. Here we report the establishment of transgenic mice in which VEGF-C expression, driven by the rat insulin promoter (Rip), is targeted to beta-cells of the endocrine pancreas. In contrast to wild-type mice, which lack peri-insular lymphatics, RipVEGF-C transgenics develop an extensive network of lymphatics around the islets of Langerhans. These mice were crossed with Rip1Tag2 mice, which develop pancreatic beta-cell tumours that are neither lymphangiogenic nor metastatic. Double-transgenic mice formed tumours surrounded by well developed lymphatics, which frequently contained tumour cell masses of beta-cell origin. These mice frequently developed pancreatic lymph node metastases. Our findings demonstrate that VEGF-C-induced lymphangiogenesis mediates tumour cell dissemination and the formation of lymph node metastases.
Figures
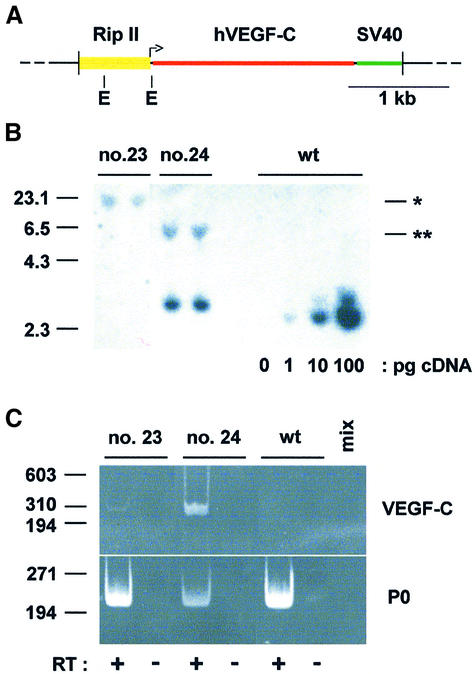
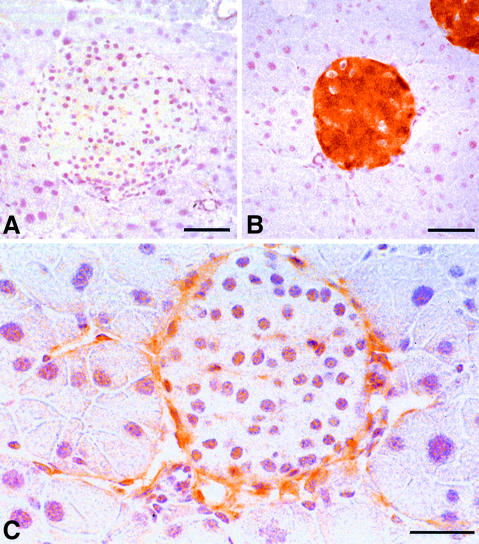
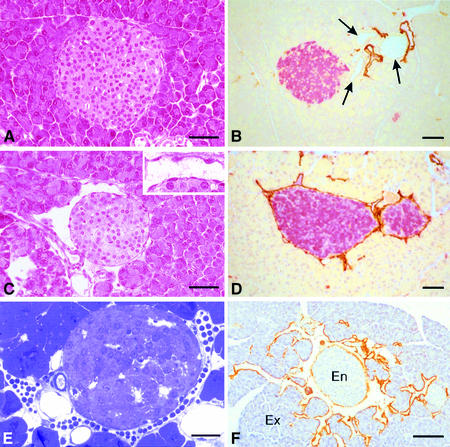
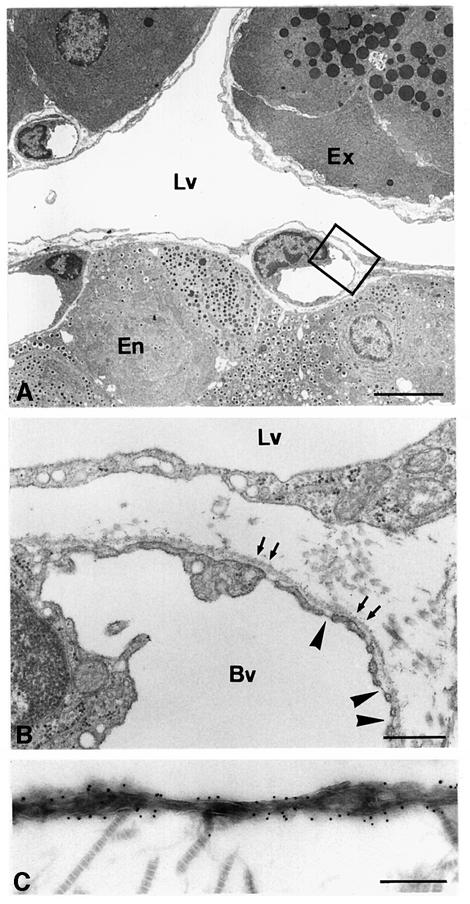
References
-
- Alpert S., Hanahan,D. and Teitelman,G. (1988) Hybrid insulin genes reveal a developmental lineage for pancreatic endocrine cells and imply a relationship with neurons. Cell, 53, 295–308. - PubMed
-
- Bertelli E., Regoli,M. and Comparini,L. (1993) Histotopographic and ultrastructural study on the lymphatic network of the pancreas in the guinea pig. Acta Anat. (Basel), 147, 233–239. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
