

The budding yeast proteins Spc24p and Spc25p interact with Ndc80p and Nuf2p at the kinetochore and are important for kinetochore clustering and checkpoint control
- PMID: 11179222
- PMCID: PMC145434
- DOI: 10.1093/emboj/20.4.777
The budding yeast proteins Spc24p and Spc25p interact with Ndc80p and Nuf2p at the kinetochore and are important for kinetochore clustering and checkpoint control
Abstract
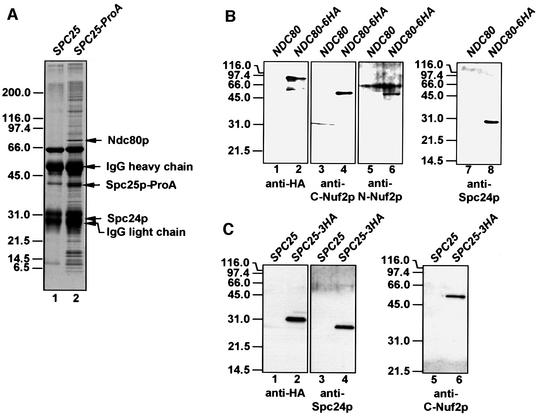
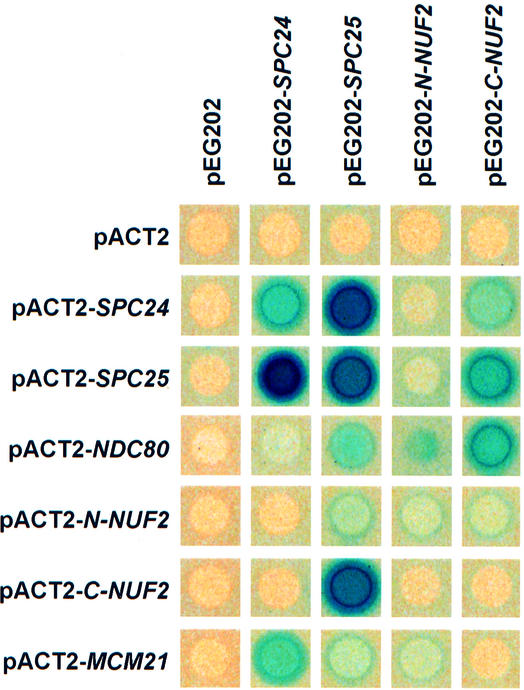
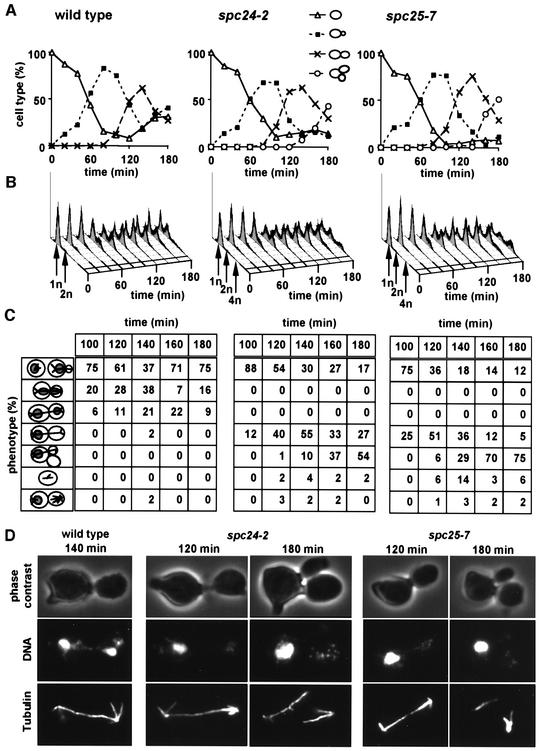
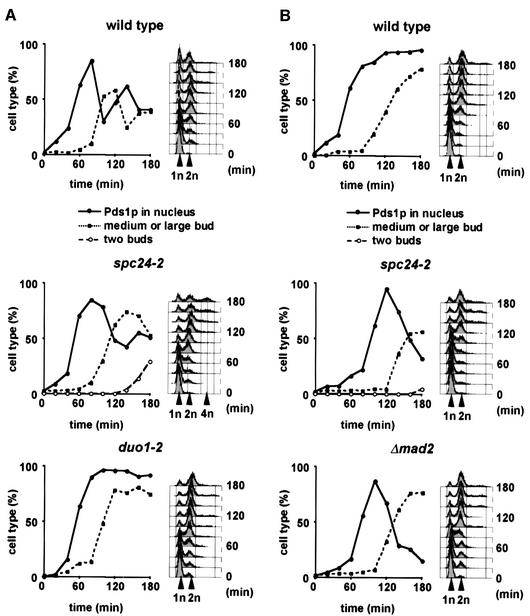
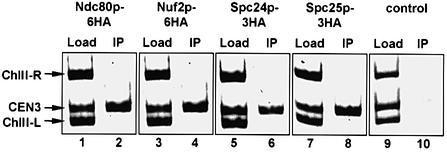
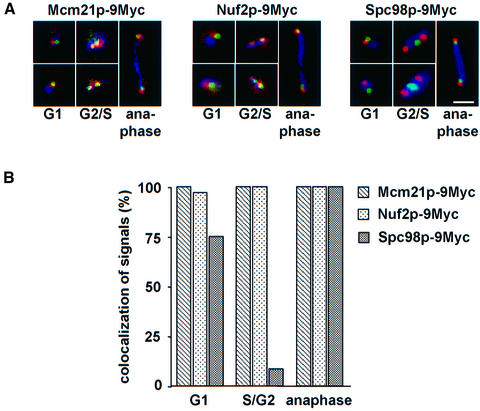
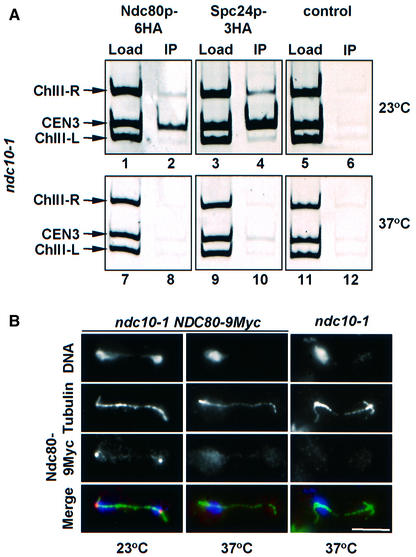
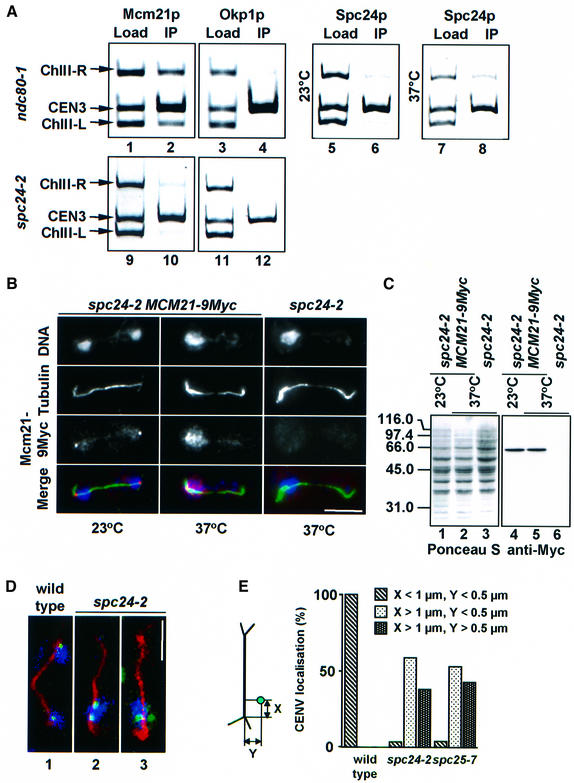
Here, we show that the budding yeast proteins Ndc80p, Nuf2p, Spc24p and Spc25p interact at the kinetochore. Consistently, Ndc80p, Nuf2p, Spc24p and Spc25p associate with centromere DNA in chromatin immunoprecipitation experiments, and SPC24 interacts genetically with MCM21 encoding a kinetochore component. Moreover, although conditional lethal spc24-2 and spc25-7 cells form a mitotic spindle, the kinetochores remain in the mother cell body and fail to segregate the chromosomes. Despite this defect in chromosome segregation, spc24-2 and spc25-7 cells do not arrest in metaphase in response to checkpoint control. Furthermore, spc24-2 cells showed a mitotic checkpoint defect when microtubules were depolymerized with nocodazole, indicating that Spc24p has a function in checkpoint control. Since Ndc80p, Nuf2p and Spc24p are conserved proteins, it is likely that similar complexes are part of the kinetochore in other organisms.
Figures
References
-
- Abrieu A., Kahana,J.A., Wood,K.W. and Cleveland,D.W. (2000) CENP-E as an essential component of the mitotic checkpoint in vitro. Cell, 102, 817–826. - PubMed
-
- Altschul S.F. and Gish,W. (1996) Local alignment statistics. Methods Enzymol., 266, 460–480. - PubMed
-
- Bardin A.J., Visintin,R. and Amon,A. (2000) A mechanism for coupling exit from mitosis to partitioning of the nucleus. Cell, 102, 21–31. - PubMed
-
- Chen R.H., Waters,J.C., Salmon,E.D. and Murray,A.W. (1996) Association of spindle assembly checkpoint component XMAD2 with unattached kinetochores. Science, 274, 242–246. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
