

The effects of pH on the interaction between capsaicin and the vanilloid receptor in rat dorsal root ganglia neurons
- PMID: 11181431
- PMCID: PMC1572632
- DOI: 10.1038/sj.bjp.0703900
The effects of pH on the interaction between capsaicin and the vanilloid receptor in rat dorsal root ganglia neurons
Abstract
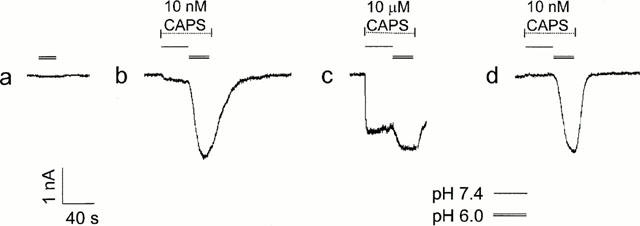
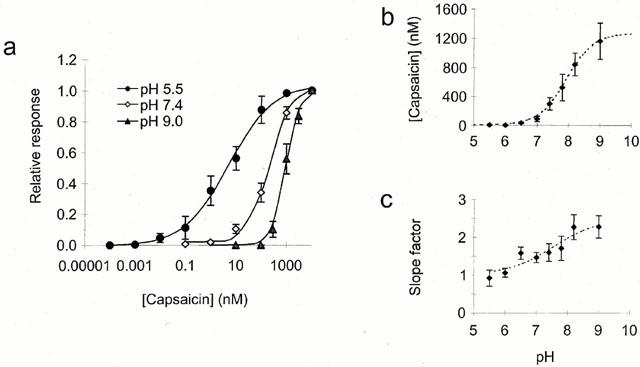
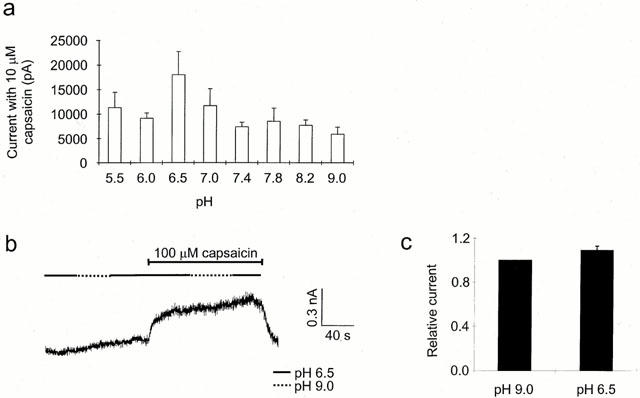
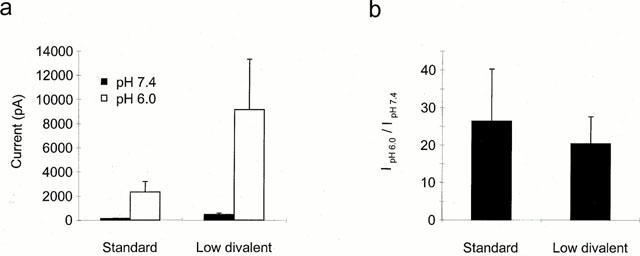
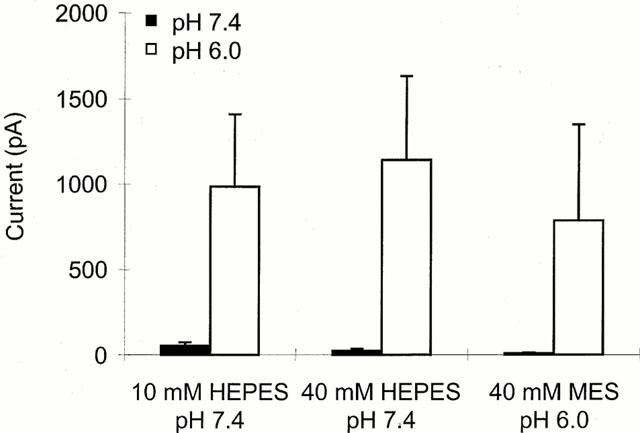
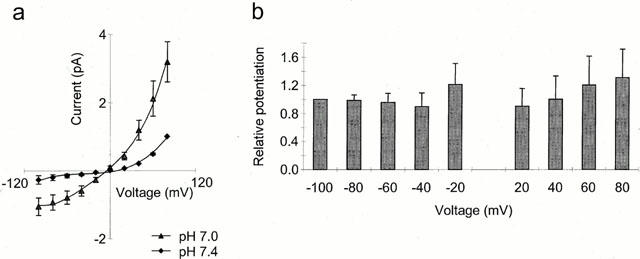
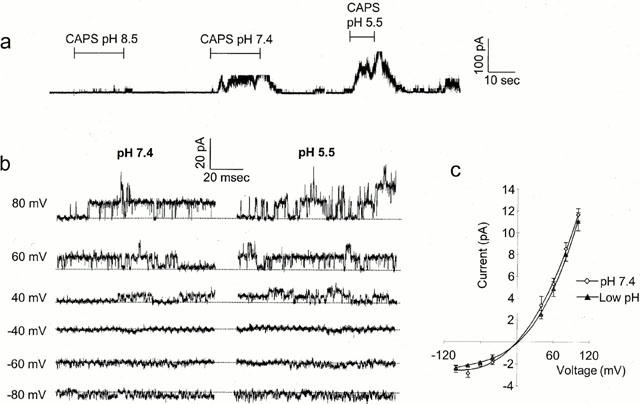
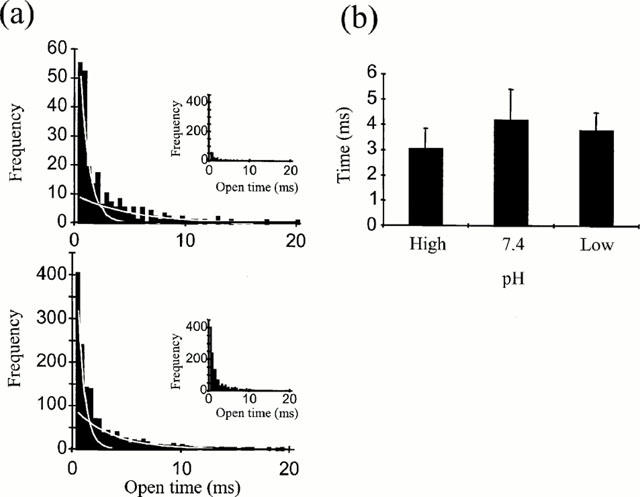
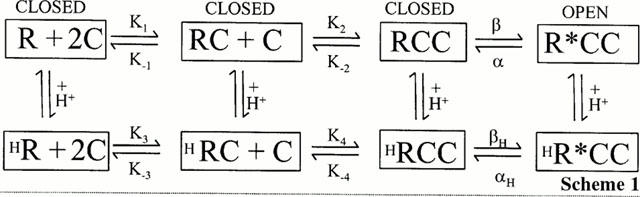
1. The vanilloid receptor of sensory neurons is a polymodal nociceptor sensitive to capsaicin, protons, heat and anandamide. Although it is known that interaction occurs between these different mediators the mechanism by which this occurs is poorly understood. In this study capsaicin elicited currents were recorded from vanilloid receptors found in adult rat isolated dorsal root ganglia (DRG) neurons under conditions of varying pH and the mechanism whereby protons can modulate this capsaicin response investigated. 2. Under whole-cell voltage clamp, modulating extracellular pH shifted the position of the capsaicin log(concentration)-response curve. Acidification from pH 9.0 to pH 5.5 lowered the EC50 values from 1150+/-250 nM to 5+/-2 nM with coincident change in the mean apparent slope factor from 2.3+/-0.3 to 0.9+/-0.2 and no change in maximal response. 3. The magnitude of the potentiation seen on reducing extracellular pH was not significantly affected by changes in extracellular calcium and magnesium concentration. 4. The response to capsaicin was not potentiated by a reduction in intracellular pH suggesting a site of action more accessible from the extracellular than the intracellular side of the membrane. 5. Potentiation by low pH was voltage independent indicating a site of action outside the membrane electric field. 6. At the single channel level, reducing extracellular pH increased channel open probability but had no significant effect on single channel conductance or open time. 7. These results are consistent with a model in which, on reducing extracellular pH, the vanilloid receptor in rat DRG neurons, changes from a state with low affinity for capsaicin to one with high affinity, coincident with a loss of cooperativity. This effect, presumed to be proton mediated, appears to involve one or more sites with pK(a) value 7.4-7.9, outside the membrane electrical field on an extracellularly exposed region of the receptor protein.
Figures
Similar articles
-
Resiniferatoxin-type phorboid vanilloids display capsaicin-like selectivity at native vanilloid receptors on rat DRG neurons and at the cloned vanilloid receptor VR1.Br J Pharmacol. 1999 Sep;128(2):428-34. doi: 10.1038/sj.bjp.0702810. Br J Pharmacol. 1999. PMID: 10510454 Free PMC article.
-
Differential pH and capsaicin responses of Griffonia simplicifolia IB4 (IB4)-positive and IB4-negative small sensory neurons.Neuroscience. 2004;127(3):659-72. doi: 10.1016/j.neuroscience.2004.05.041. Neuroscience. 2004. PMID: 15283965
-
Protein kinase C activation potentiates gating of the vanilloid receptor VR1 by capsaicin, protons, heat and anandamide.J Physiol. 2001 Aug 1;534(Pt 3):813-25. doi: 10.1111/j.1469-7793.2001.00813.x. J Physiol. 2001. PMID: 11483711 Free PMC article.
-
Recent advances in understanding of vanilloid receptors: a therapeutic target for treatment of pain and inflammation in skin.J Investig Dermatol Symp Proc. 1997 Aug;2(1):56-60. doi: 10.1038/jidsymp.1997.12. J Investig Dermatol Symp Proc. 1997. PMID: 9487017 Review.
-
The vanilloid (capsaicin) receptor: receptor types and species differences.Gen Pharmacol. 1994 Mar;25(2):223-43. doi: 10.1016/0306-3623(94)90049-3. Gen Pharmacol. 1994. PMID: 8026721 Review.
Cited by
-
Functional characterisation of the S512Y mutant vanilloid human TRPV1 receptor.Br J Pharmacol. 2005 Nov;146(5):702-11. doi: 10.1038/sj.bjp.0706356. Br J Pharmacol. 2005. PMID: 16100528 Free PMC article.
-
Capsaicin, a Powerful •OH-Inactivating Ligand.Antioxidants (Basel). 2020 Dec 8;9(12):1247. doi: 10.3390/antiox9121247. Antioxidants (Basel). 2020. PMID: 33302572 Free PMC article.
-
TRPV1 (vanilloid receptor) in the urinary tract: expression, function and clinical applications.Naunyn Schmiedebergs Arch Pharmacol. 2006 Jul;373(4):287-99. doi: 10.1007/s00210-006-0073-2. Epub 2006 May 24. Naunyn Schmiedebergs Arch Pharmacol. 2006. PMID: 16721555 Review.
-
'Entourage' effects of N-acyl ethanolamines at human vanilloid receptors. Comparison of effects upon anandamide-induced vanilloid receptor activation and upon anandamide metabolism.Br J Pharmacol. 2002 Jun;136(3):452-8. doi: 10.1038/sj.bjp.0704732. Br J Pharmacol. 2002. PMID: 12023948 Free PMC article.
-
The pharmacological challenge to tame the transient receptor potential vanilloid-1 (TRPV1) nocisensor.Br J Pharmacol. 2008 Dec;155(8):1145-62. doi: 10.1038/bjp.2008.351. Epub 2008 Sep 22. Br J Pharmacol. 2008. PMID: 18806809 Free PMC article. Review.
References
-
- BEVAN S., DOCHERTY R.J.Cellular mechanisms of the action of capsaicin Capsaicin in the study of pain 1993Academic Press; 27–44.ed. Wood, J. Chapter 2, pp
-
- BEVAN S., FORBES C.A., WINTER J. Protons and capsaicin activate the same ion channels in rat isolated dorsal root ganglion cells. J. Physiol. 1993;459:401P.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
