

Increased atherosclerosis in myeloperoxidase-deficient mice
- PMID: 11181641
- PMCID: PMC199241
- DOI: 10.1172/JCI8797
Increased atherosclerosis in myeloperoxidase-deficient mice
Abstract
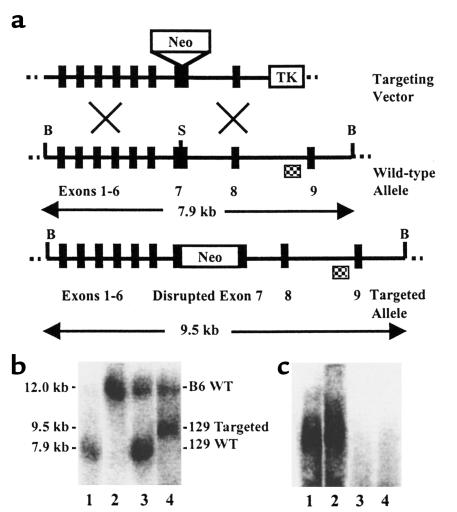
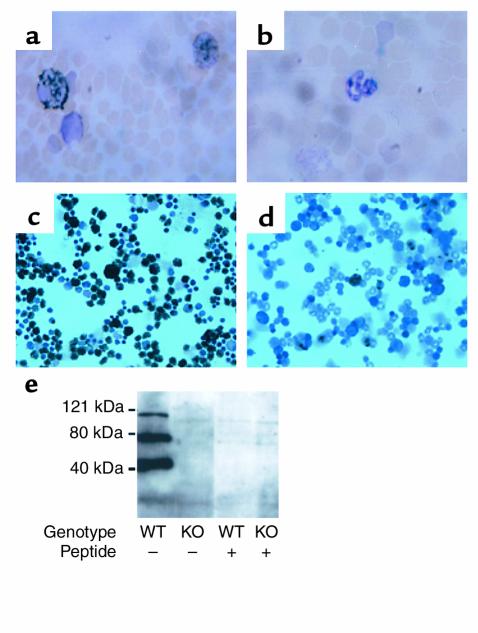
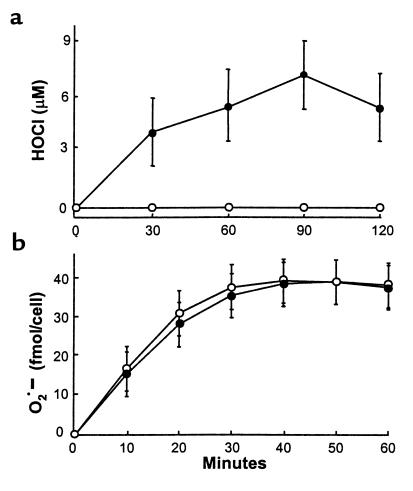
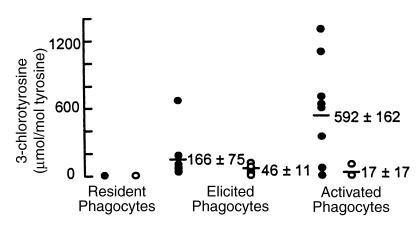
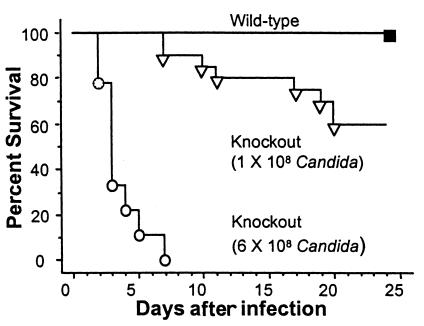
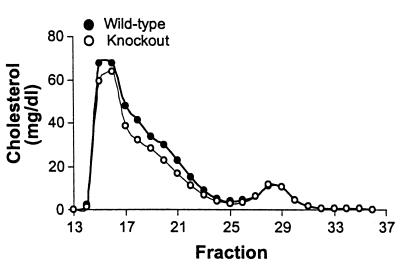
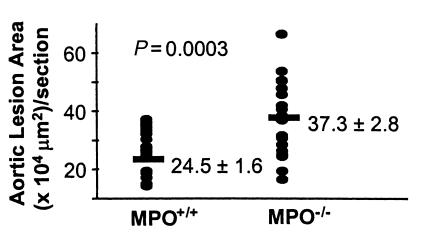
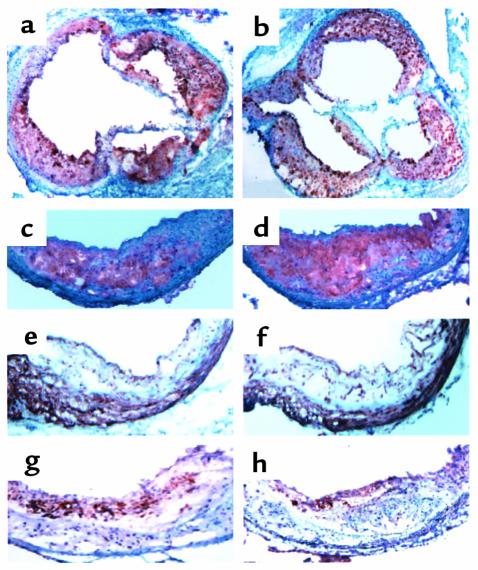
Myeloperoxidase (MPO), a heme enzyme secreted by activated phagocytes, generates an array of oxidants proposed to play critical roles in host defense and local tissue damage. Both MPO and its reaction products are present in human atherosclerotic plaque, and it has been proposed that MPO oxidatively modifies targets in the artery wall. We have now generated MPO-deficient mice, and show here that neutrophils from homozygous mutants lack peroxidase and chlorination activity in vitro and fail to generate chlorotyrosine or to kill Candida albicans in vivo. To examine the potential role of MPO in atherosclerosis, we subjected LDL receptor-deficient mice to lethal irradiation, repopulated their marrow with MPO-deficient or wild-type cells, and provided them a high-fat, high-cholesterol diet for 14 weeks. White cell counts and plasma lipoprotein profiles were similar between the two groups at sacrifice. Cross-sectional analysis of the aorta indicated that lesions in MPO-deficient mice were about 50% larger than controls. Similar results were obtained in a genetic cross with LDL receptor-deficient mice. In contrast to advanced human atherosclerotic lesions, the chlorotyrosine content of aortic lesions from wild-type as well as MPO-deficient mice was essentially undetectable. These data suggest an unexpected, protective role for MPO-generated reactive intermediates in murine atherosclerosis. They also identify an important distinction between murine and human atherosclerosis with regard to the potential involvement of MPO in protein oxidation.
Figures
Comment in
-
The proper study of mankind.J Clin Invest. 2001 Feb;107(4):401-3. doi: 10.1172/JCI11713. J Clin Invest. 2001. PMID: 11181638 Free PMC article. No abstract available.
References
-
- Agner K. Verdoperoxidase: a ferment isolated from leukocytes. Acta Physiol Scand. 1941; 2:1–62.
-
- Babior BM. Oxygen-dependent microbial killing by phagocytes. N Engl J Med. 1978; 298:659–666. - PubMed
-
- Klebanoff, S.J., and Clark, R.A. 1978. The neutrophil: function and clinical disorders. Elsevier/North Holland Biomedical Press. Amsterdam, The Netherlands. 1–810.
-
- Weiss SJ. Tissue destruction by neutrophils. N Engl J Med. 1989; 320:365–376. - PubMed
-
- Shultz J, Kaminker K. Myeloperoxidase of the leucocyte of normal human blood. 1. Content and localization. Arch Biochem Biophys. 1962; 96:465–467. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
