

Molecular and physiological diversity of nicotinic acetylcholine receptors in the midbrain dopaminergic nuclei
- PMID: 11222635
- PMCID: PMC6762941
- DOI: 10.1523/JNEUROSCI.21-05-01452.2001
Molecular and physiological diversity of nicotinic acetylcholine receptors in the midbrain dopaminergic nuclei
Abstract
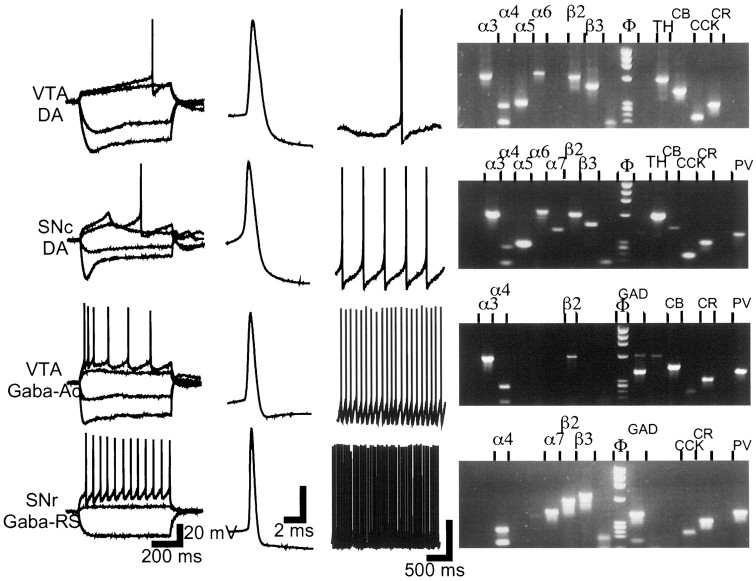
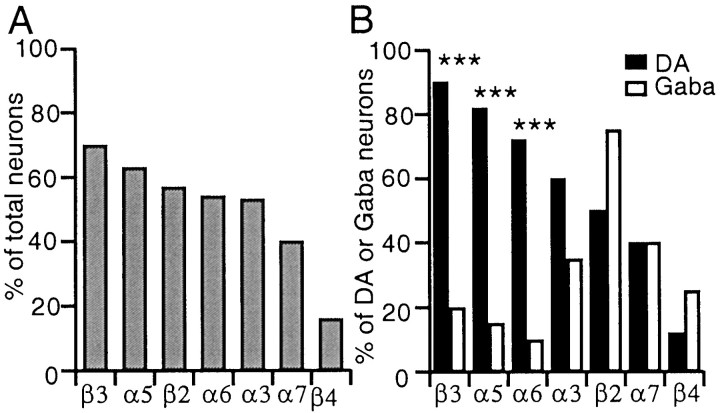
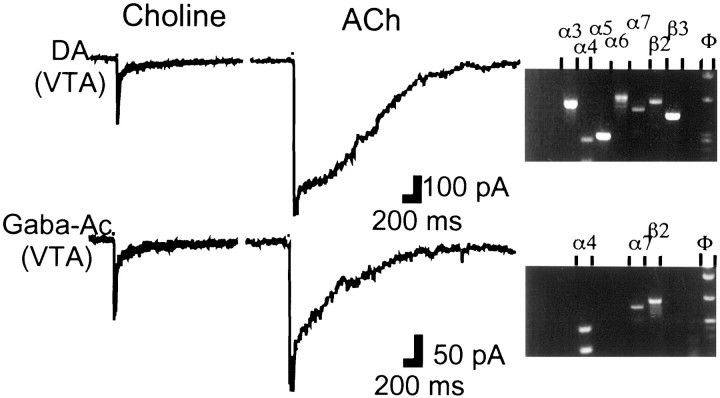
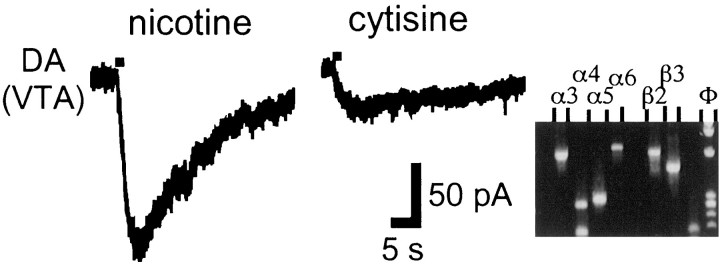
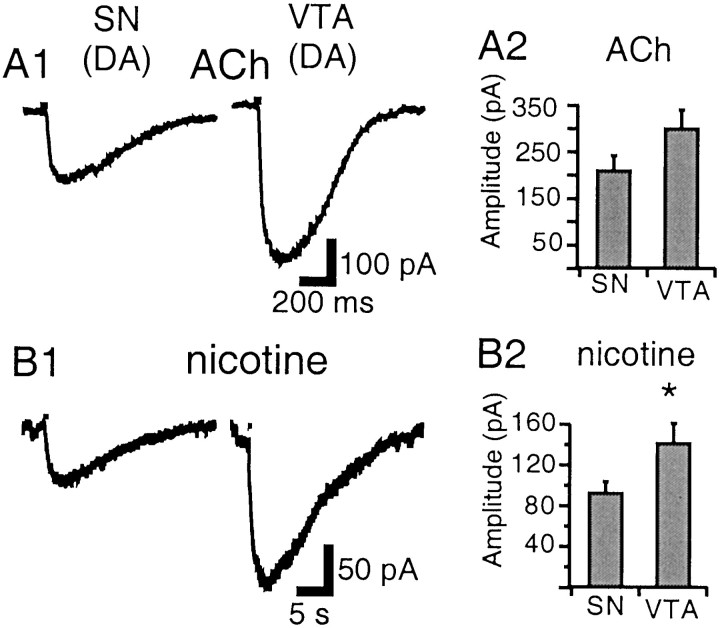
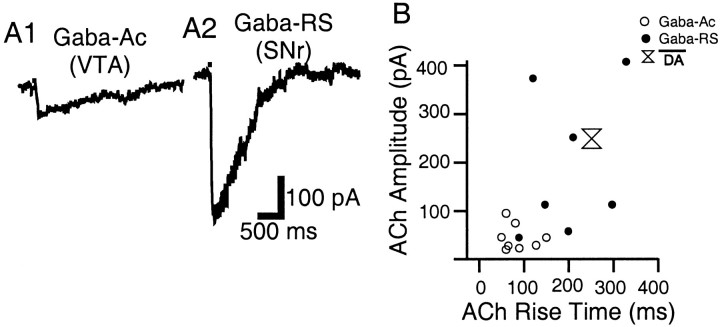
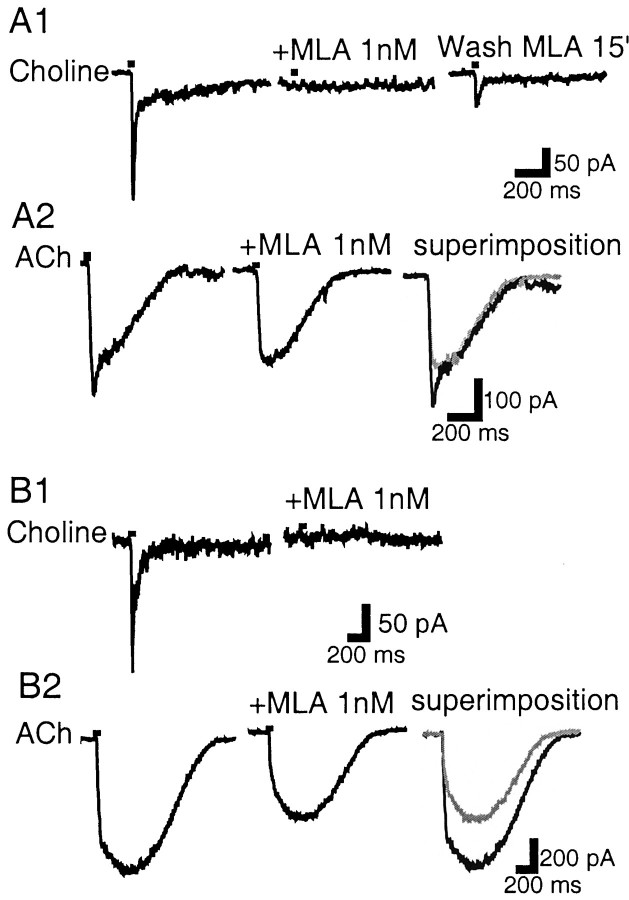
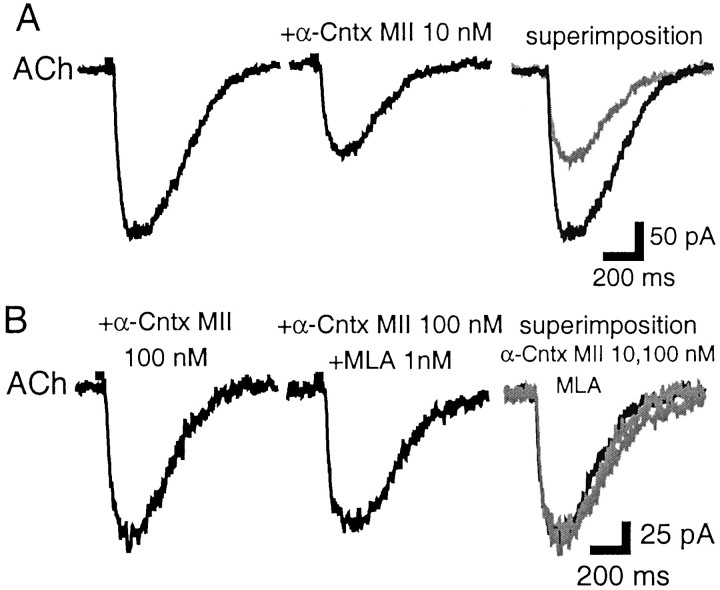
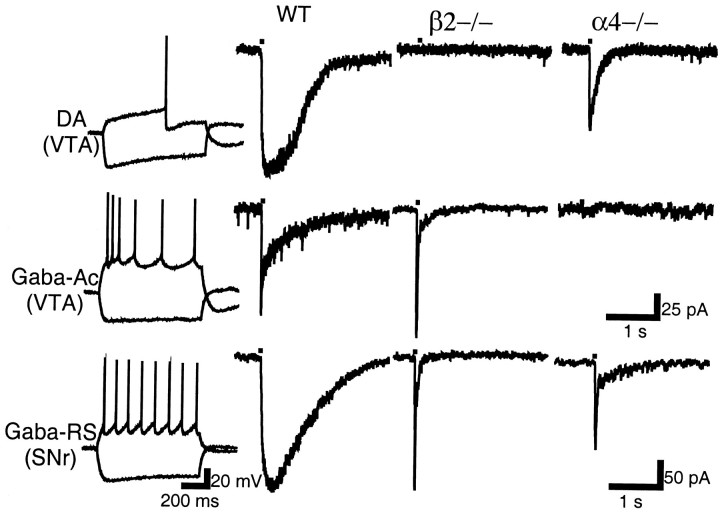
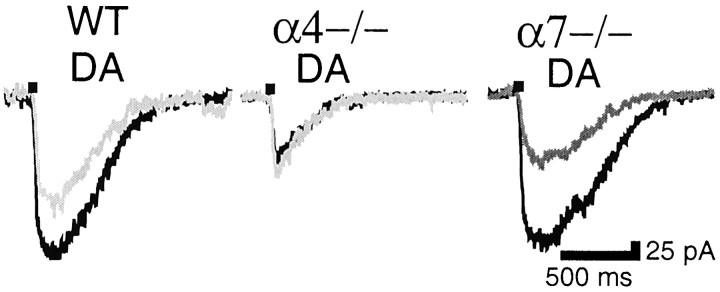
Nicotinic acetylcholine receptors (nAChRs) on dopaminergic (DA) and GABAergic (Gaba) projection neurons of the substantia nigra (SN) and ventral tegmental area (VTA) are characterized by single-cell RT-PCR and patch-clamp recordings in slices of rat and wild-type, beta2-/-, alpha4-/-, and alpha7-/- mice. The eight nAChR subunits expressed in these nuclei, alpha3-7 and beta2-4, contribute to four different types of nAChR-mediated currents. Most DA neurons in the SN and VTA express two nAChR subtypes. One is inhibited by dihydro-beta-erythroidine (2 microm), alpha-conotoxin MII (10 nm), and methyllycaconitine (1 nm) but does not contain the alpha7 subunit; it possesses a putative alpha4alpha6alpha5(beta2)(2) composition. The other subtype is inhibited by dihydro-beta-erythroidine (2 microm) and has a putative alpha4alpha5(beta2)(2) composition. Gaba neurons in the VTA exhibit a third subtype with a putative (alpha4)(2)(beta2)(3) composition, whereas Gaba neurons in the SN have either the putative (alpha4)(2)(beta2)(3) oligomer or the putative alpha4alpha6alpha5(beta2)(2) oligomer. The fourth subtype, a putative (alpha7)(5) homomer, is encountered in less than half of DA and Gaba neurons, in the SN as well as in the VTA. Neurons in the DA nuclei thus exhibit a diversity of nAChRs that might differentially modulate reinforcement and motor behavior.
Figures

References
-
- Alkondon M, Albuquerque EX. Diversity of nicotinic acetylcholine receptors in rat hippocampal neurons. I. Pharmacological and functional evidence for distinct structural subtypes. J Pharmacol Exp Ther. 1993;265:1455–1473. - PubMed
-
- Alkondon M, Pereira EF, Cortes WS, Maelicke A, Albuquerque EX. Choline is a selective agonist of α7 nicotinic acetylcholine receptors in the rat brain neurons. Eur J Neurosci. 1997;9:2734–2742. - PubMed
-
- Arroyo-Jiménez MM. PhD thesis. Universidad Miguel Hernández, Albacete, Spain; 2000. Localization of the α4 and β3 subunits of the neuronal nicotinic acetylcholine receptor in the rodent central nervous system.
-
- Baron JA. Cigarette smoking and Parkinson's disease. Neurology. 1986;36:1490–1496. - PubMed
-
- Berke JD, Hyman SE. Addiction, dopamine, and the molecular mechanisms of memory. Neuron. 2000;25:515–532. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases