

Increased expression of MIP-1 alpha and MIP-1 beta mRNAs in the brain correlates spatially and temporally with the spongiform neurodegeneration induced by a murine oncornavirus
- PMID: 11222690
- PMCID: PMC115891
- DOI: 10.1128/JVI.75.6.2665-2674.2001
Increased expression of MIP-1 alpha and MIP-1 beta mRNAs in the brain correlates spatially and temporally with the spongiform neurodegeneration induced by a murine oncornavirus
Abstract
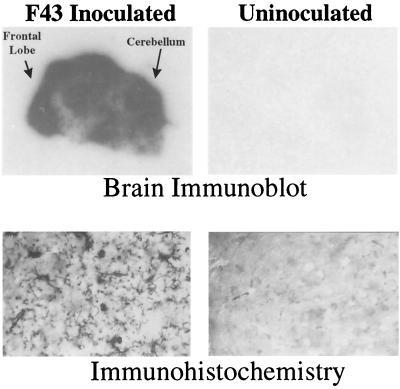
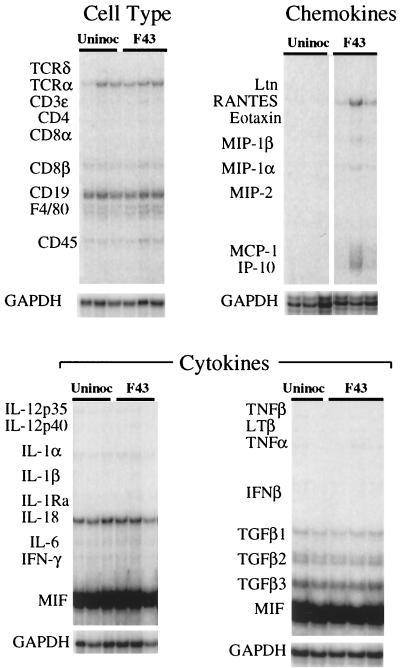
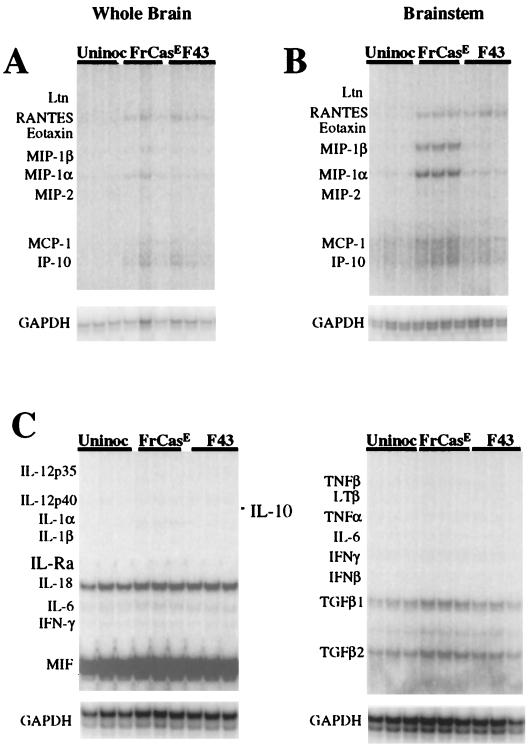
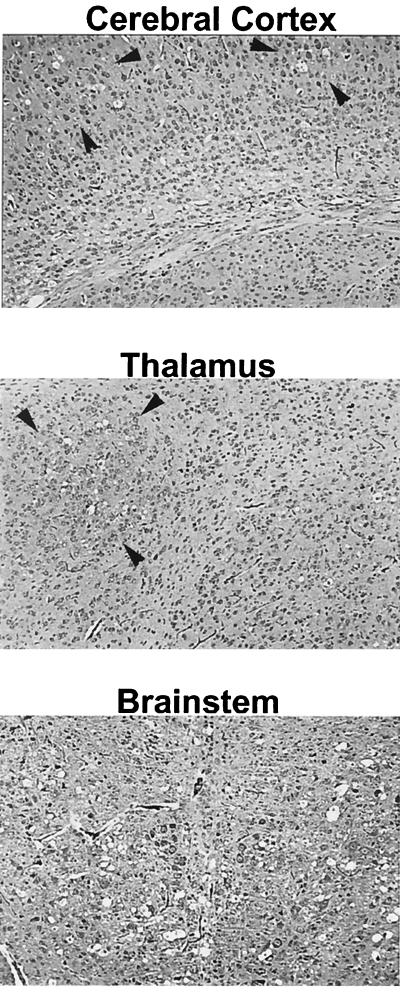
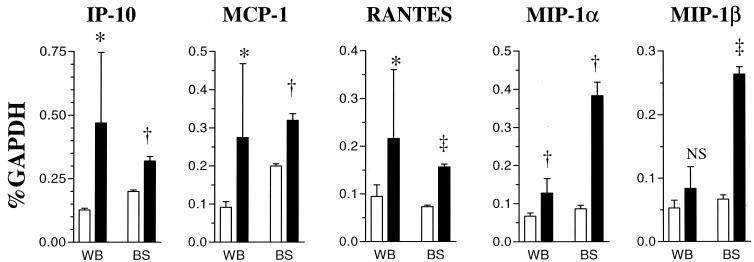
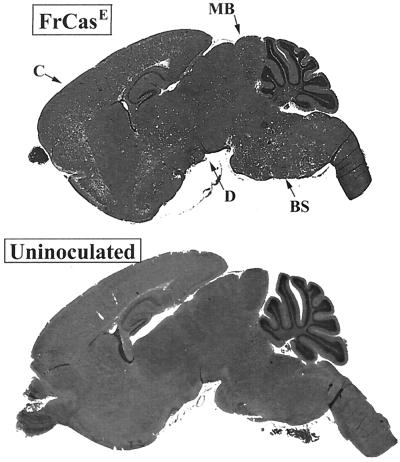
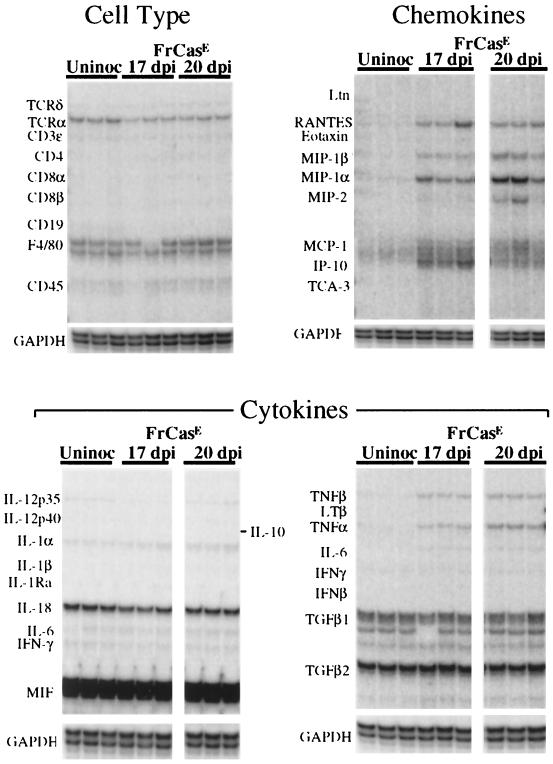
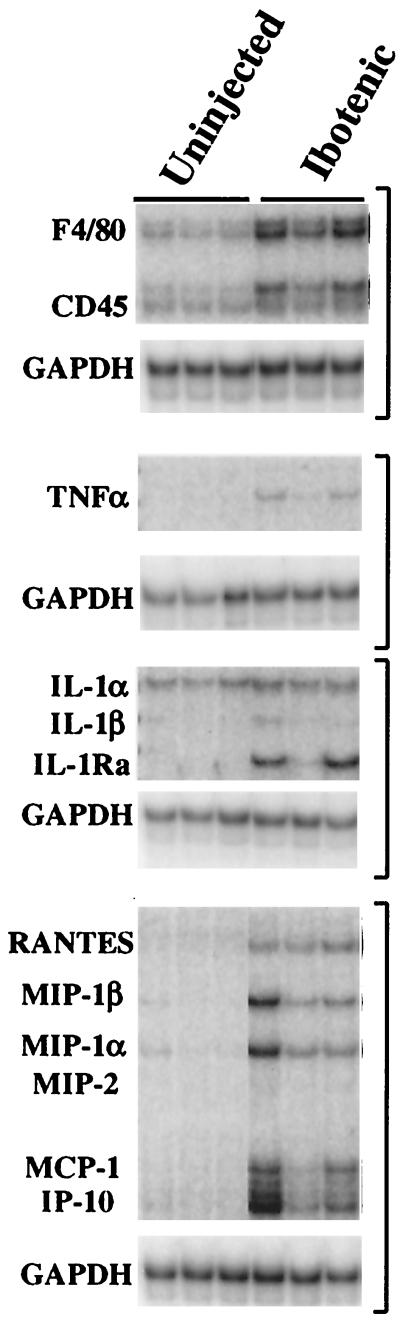
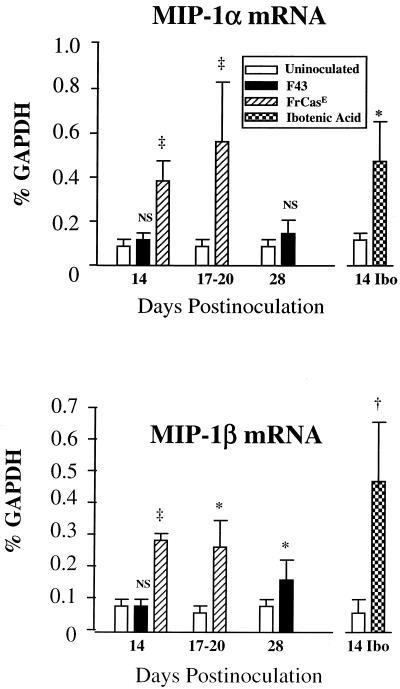
The chimeric murine oncornavirus FrCas(E) causes a rapidly progressive paralytic disease associated with spongiform neurodegeneration throughout the neuroaxis. Neurovirulence is determined by the sequence of the viral envelope gene and by the capacity of the virus to infect microglia. The neurocytopathic effect of this virus appears to be indirect, since the cells which degenerate are not infected. In the present study we have examined the possible role of inflammatory responses in this disease and have used as a control the virus F43. F43 is an highly neuroinvasive but avirulent virus which differs from FrCas(E) only in 3' pol and env sequences. Like FrCas(E), F43 infects large numbers of microglial cells, but it does not induce spongiform neurodegeneration. RNAase protection assays were used to detect differential expression of genes encoding a variety of cytokines, chemokines, and inflammatory cell-specific markers. Tumor necrosis factor alpha (TNF-alpha) and TNF-beta mRNAs were upregulated in advanced stages of disease but not early, even in regions with prominent spongiosis. Surprisingly there was no evidence for upregulation of the cytokines interleukin-1 alpha (IL-1 alpha), IL-1 beta, and IL-6 or of the microglial marker F4/80 at any stage of this disease. In contrast, increased levels of the beta-chemokines MIP-1 alpha and -beta were seen early in the disease and were concentrated in regions of the brain rich in spongiosis, and the magnitude of responses was similar to that observed in the brains of mice injected with the glutamatergic neurotoxin ibotenic acid. MIP-1alpha and MIP-1beta mRNAs were also upregulated in F43-inoculated mice, but the responses were three- to fivefold lower and occurred later in the course of infection than was observed in FrCas(E)-inoculated mice. These results suggest that the robust increase in expression of MIP-1 alpha and MIP-1 beta in the brain represents a correlate of neurovirulence in this disease, whereas the TNF responses are likely secondary events.
Figures
References
-
- Andrews J M, Gardner M B. Lower motor neuron degeneration associated with type C RNA virus infection in mice: neuropathological features. J Neuropathol Exp Neurol. 1974;33:285–307. - PubMed
-
- Asensio V C, Kincaid C, Campbell I L. Chemokines and the inflammatory response to viral infection in the central nervous system with a focus on lymphocytic choriomeningitis virus. J Neurovirol. 1999;5:65–75. - PubMed
-
- Bell M D, Taub D D, Perry V H. Overriding the brain's intrinsic resistance to leukocyte recruitment with intraparenchymal injections of recombinant chemokines. Neuroscience. 1996;74:283–292. - PubMed
-
- Betmouni S, Perry V H, Gordon J L. Evidence for an early inflammatory response in the central nervous system of mice with scrapie. Neuroscience. 1996;74:1–5. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
