

Polysomal ribonuclease 1 exists in a latent form on polysomes prior to estrogen activation of mRNA decay
- PMID: 11222765
- PMCID: PMC29717
- DOI: 10.1093/nar/29.5.1156
Polysomal ribonuclease 1 exists in a latent form on polysomes prior to estrogen activation of mRNA decay
Abstract
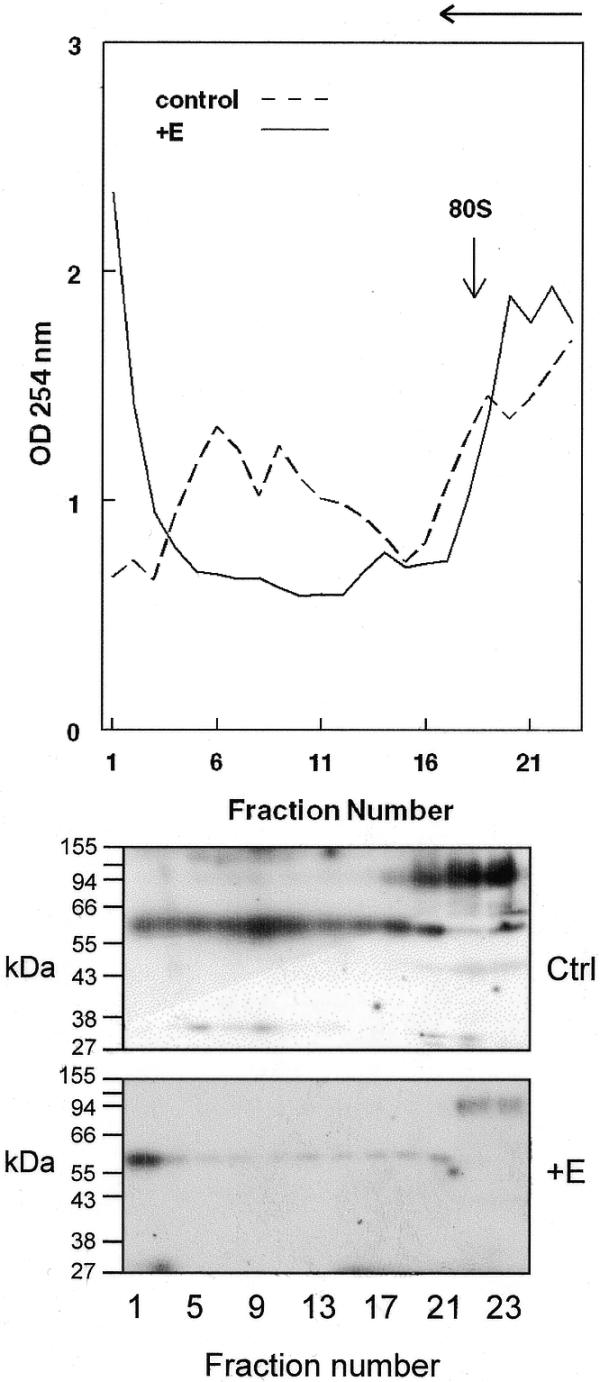
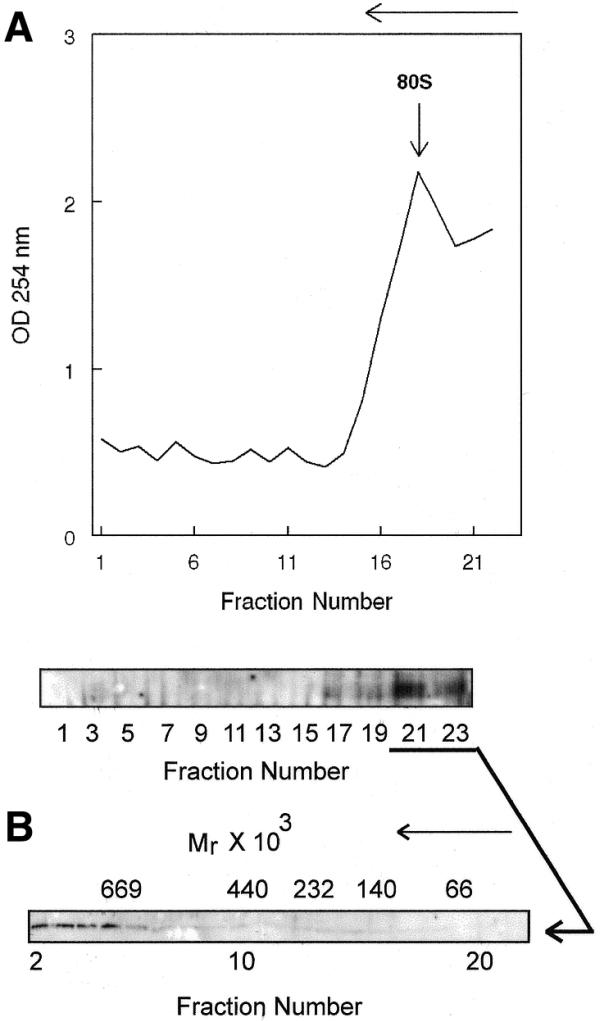
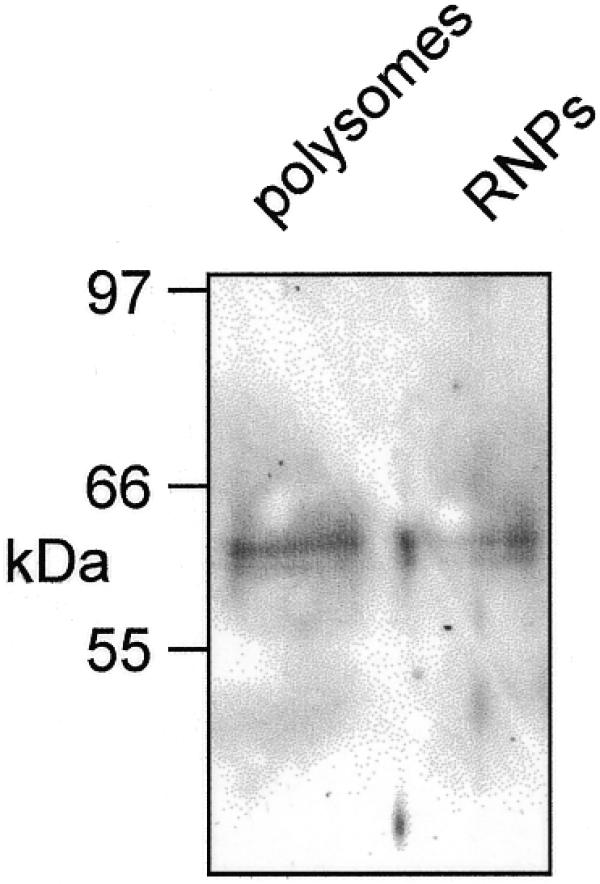
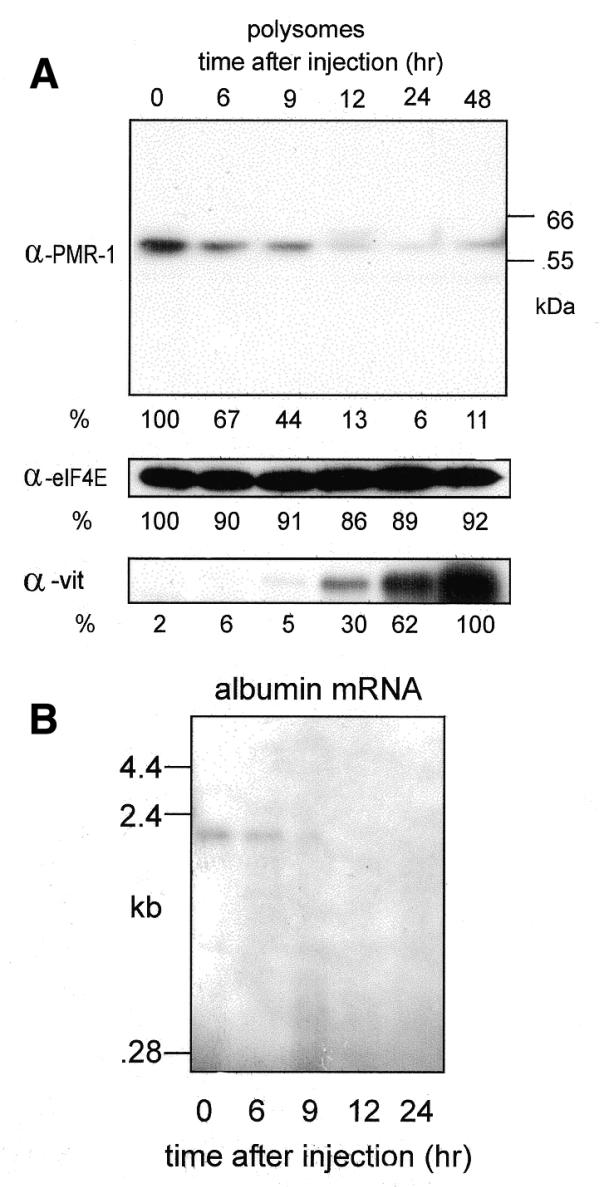
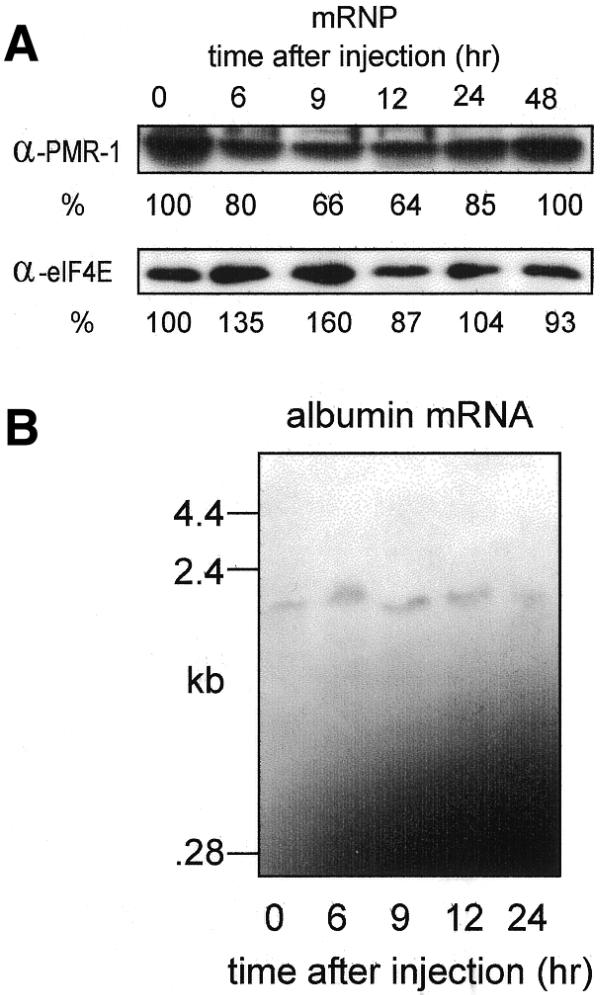
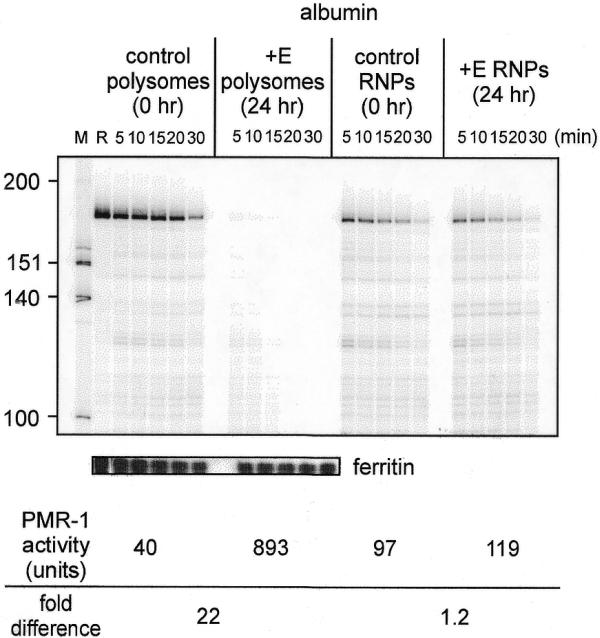
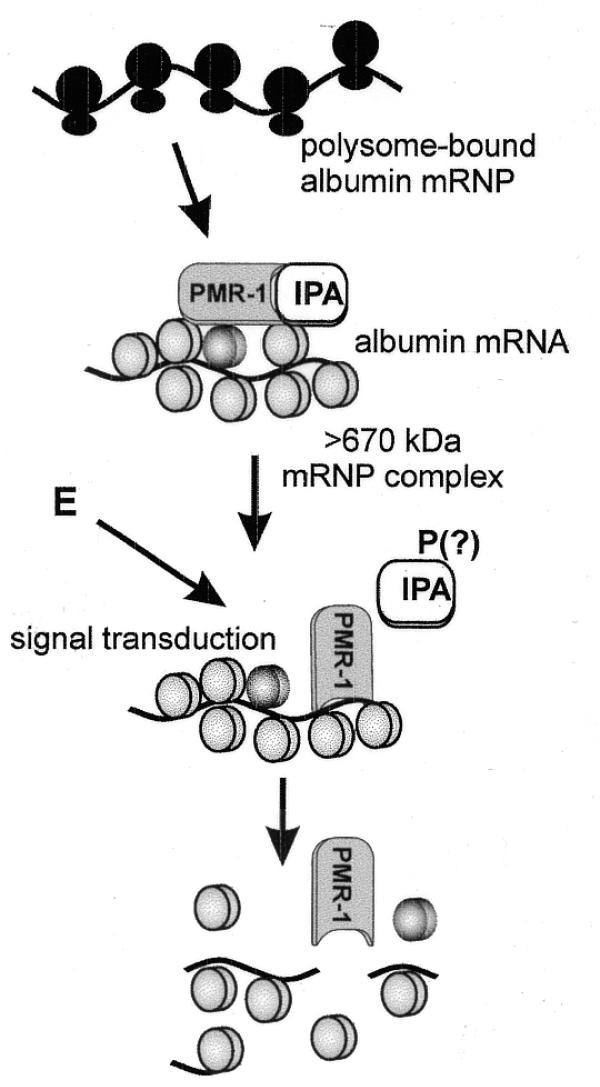
Estrogen induces a global change in the translation profile of Xenopus hepatocytes, replacing serum protein synthesis with production of the yolk protein precursor vitellogenin. This is accomplished by the coordinate destabilization of serum protein mRNAs and the transcriptional induction and subsequent stabilization of vitellogenin mRNA. Previous work identified an endonuclease activity whose appearance on polysomes correlated with the disappearance of serum protein mRNAs. This enzyme, polysomal ribonuclease 1 (PMR1), is a novel member of the peroxidase gene family. The current study examined the association of PMR1 with its mRNA targets on polysomes and mRNPs. The highest amount of polysome-bound PMR1 was observed prior to estrogen induction of mRNA decay. Its distribution on sucrose density gradients matched the absorbance profile of polysome-bound mRNA, suggesting that PMR1 forms a latent complex with mRNA. Following dissociation with EDTA the 62 kDa PMR1 sedimented with a larger complex of >670 kDa. Estrogen induces a 22-fold increase in unit enzymatic activity of polysome-bound PMR1, and a time-dependent loss of PMR1 from polysomes in a manner that mirrors the disappearance of albumin mRNA. These data suggest that the key step in the extensive estrogen-induced change in mRNA decay in Xenopus liver is activation of a latent mRNA endonuclease associated with its target mRNA.
Figures
References
-
- Ross J. (1996) Control of messenger RNA stability in higher eukaryotes. Trends. Genet., 12, 171–175. - PubMed
-
- Jacobson A. and Peltz,S.W. (1996) Interrelationships of the pathways of mRNA decay and translation in eukaryotic cells. Annu. Rev. Biochem., 65, 693–739. - PubMed
-
- Tharun S. and Parker,R. (1997) Mechanisms of mRNA turnover in eukaryotic cells. In Harford,J. and Morris,D.R. (eds), mRNA Metabolism and Post-transcriptional Gene Regulation. Wiley, New York, NY, pp. 181–200.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
