

Coiled-coil domain-mediated FRQ-FRQ interaction is essential for its circadian clock function in Neurospora
- PMID: 11226160
- PMCID: PMC140186
- DOI: 10.1093/emboj/20.1.101
Coiled-coil domain-mediated FRQ-FRQ interaction is essential for its circadian clock function in Neurospora
Abstract
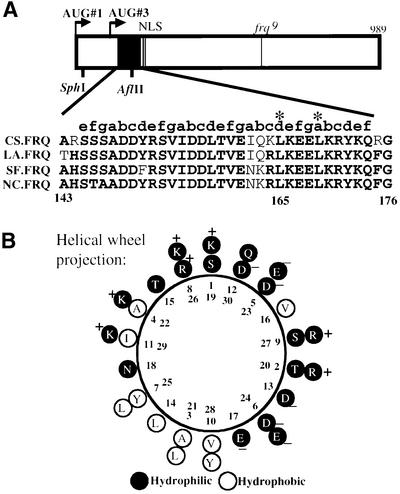
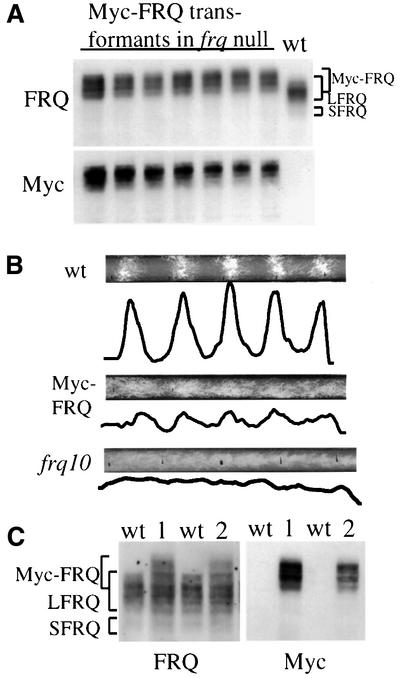
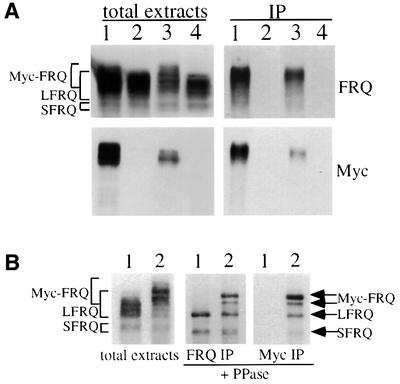
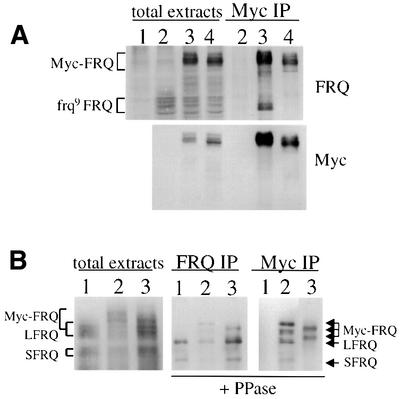
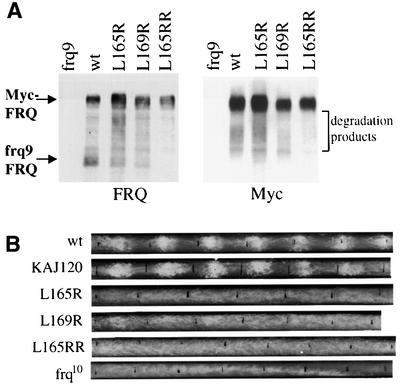
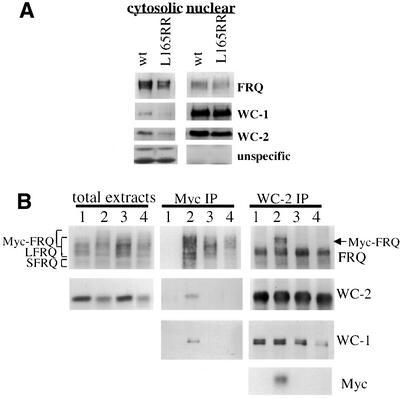
The frequency (frq) gene, the central component of the frq-based circadian negative feedback loop, regulates various aspects of the circadian clock in NEUROSPORA: However, the biochemical function of its protein products, FRQ, is poorly understood. In this study, we demonstrated that the most conserved region of FRQ forms a coiled-coil domain. FRQ interacts with itself in vivo, and the deletion of the coiled-coil region results in loss of the interaction. Point mutations, which are designed to disrupt the coiled-coil structure, weaken or completely abolish the FRQ self-association and lead to the arrhythmicity of the overt rhythm. Mutations of the FRQ coiled-coil that inhibit self-association also prevent its interaction with two other key components of the NEUROSPORA: circadian clock, namely WC-1 and WC-2, the two PAS domain-containing transcription factors. Taken together, these data strongly suggest that the formation of the FRQ-FRQ and FRQ-WC complexes is essential for the function of the NEUROSPORA: clock.
Figures
References
-
- Allada R., White,N.E., So,W.V., Hall,J.C. and Rosbash,M. (1998) A mutant Drosophila homolog of mammalian Clock disrupts circadian rhythms and transcription of period and timeless. Cell, 93, 791–804. - PubMed
-
- Aronson B., Johnson,K., Loros,J.J. and Dunlap,J.C. (1994a) Negative feedback defining a circadian clock: autoregulation in the clock gene frequency. Science, 263, 1578–1584. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
