

A cytosolic NAD-dependent deacetylase, Hst2p, can modulate nucleolar and telomeric silencing in yeast
- PMID: 11226170
- PMCID: PMC140183
- DOI: 10.1093/emboj/20.1.197
A cytosolic NAD-dependent deacetylase, Hst2p, can modulate nucleolar and telomeric silencing in yeast
Abstract
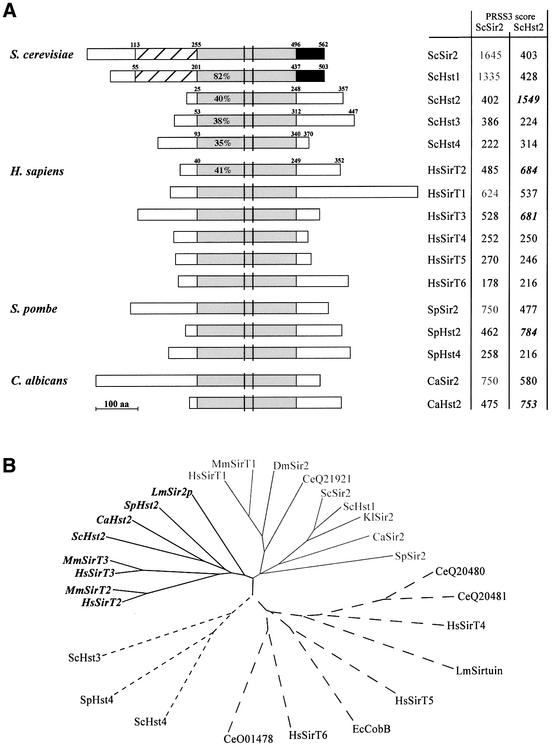
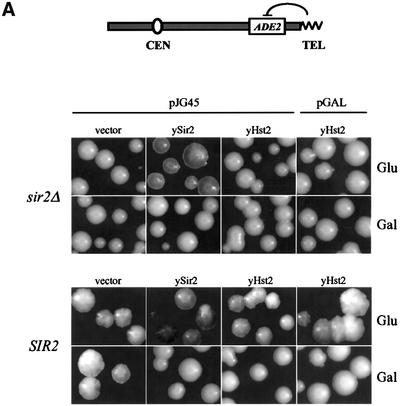
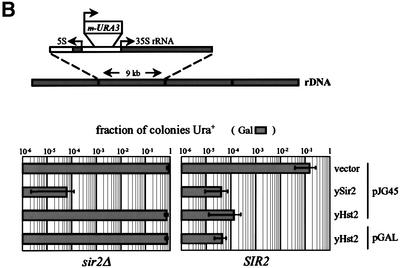
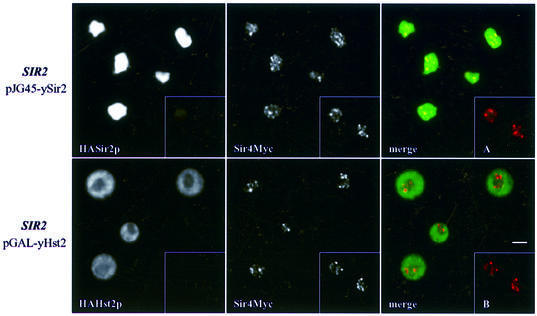
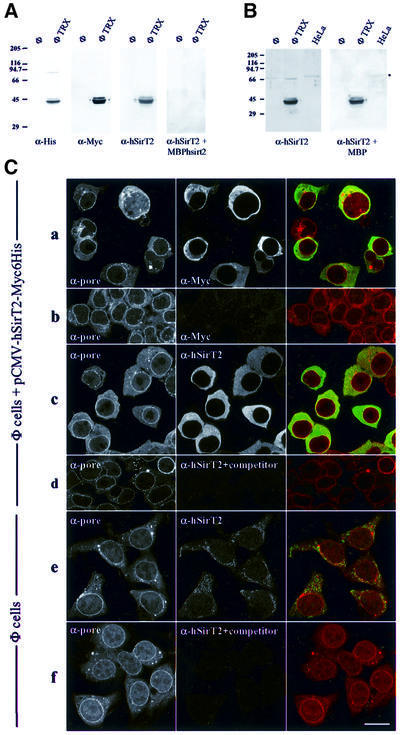
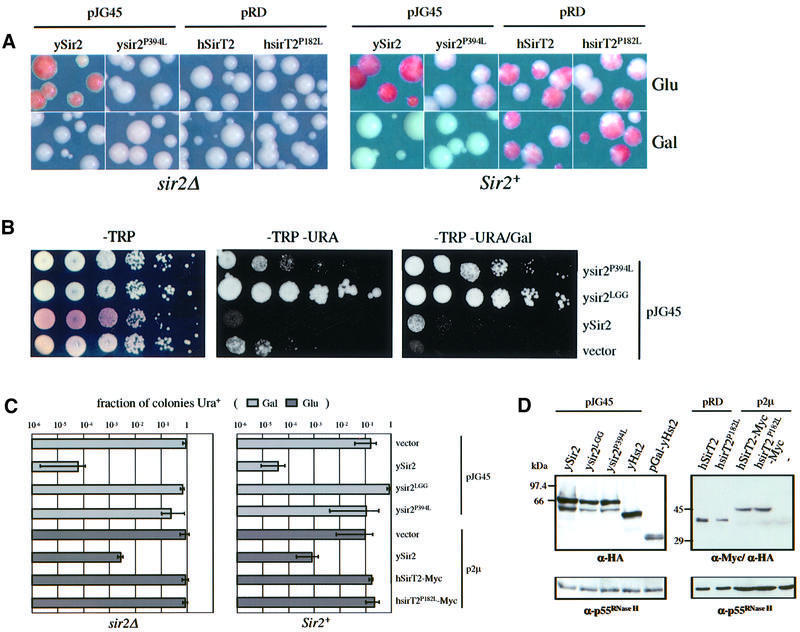
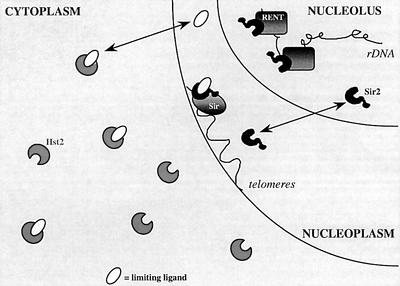
In budding yeast, the silent information regulator Sir2p is a nuclear NAD-dependent deacetylase that is essential for both telomeric and rDNA silencing. All eukaryotic species examined to date have multiple homologues of Sir two (HSTs), which share a highly conserved globular core domain. Here we report that yeast Hst2p and a mammalian Hst2p homologue, hSirT2p, are cytoplasmic in yeast and human cells, in contrast to yHst1p and ySir2p which are exclusively nuclear. Although yHst2p cannot restore silencing in a sir2 deletion, overexpression of yHst2p influences nuclear silencing events in a SIR2 strain, derepressing subtelomeric silencing while increasing repression in the rDNA. In contrast, a form of ySir2p carrying a point mutation in the conserved core domain disrupts both telomeric position effect (TPE) and rDNA repression at low expression levels. This argues that non-nuclear yHst2p can compete for a substrate or ligand specifically required for telomeric, and not rDNA repression.
Figures

References
-
- Afshar G. and Murnane,J.P. (1999) Characterization of a human gene with sequence homology to Saccharomyces cerevisiae SIR2. Gene, 234, 161–168. - PubMed
-
- Aparicio O.M., Billington,B.L. and Gottschling,D.E. (1991) Modifiers of position effect are shared between telomeric and silent mating-type loci in S.cerevisiae. Cell, 66, 1279–1287. - PubMed
-
- Brachmann C.B., Sherman,J.M., Devine,S.E., Cameron,E.E., Pillus,L. and Boeke,J.D. (1995) The SIR2 gene family, conserved from bacteria to humans, functions in silencing, cell cycle progression and chromosome stability. Genes Dev., 9, 2888–2902. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
