

doi: 10.1093/emboj/20.1.285.
The SurA periplasmic PPIase lacking its parvulin domains functions in vivo and has chaperone activity
Affiliations
- PMID: 11226178
- PMCID: PMC140197
- DOI: 10.1093/emboj/20.1.285
Item in Clipboard
The SurA periplasmic PPIase lacking its parvulin domains functions in vivo and has chaperone activity
EMBO J.
.
Abstract
The Escherichia coli periplasmic peptidyl-prolyl isomerase (PPIase) SurA is involved in the maturation of outer membrane porins. SurA consists of a substantial N-terminal region, two iterative parvulin-like domains and a C-terminal tail. Here we show that a variant of SurA lacking both parvulin-like domains exhibits a PPIase-independent chaperone-like activity in vitro and almost completely complements the in vivo function of intact SurA. SurA interacts preferentially (>50-fold) with in vitro synthesized porins over other similarly sized proteins, leading us to suggest that the chaperone-like function of SurA preferentially facilitates maturation of outer membrane proteins.
Figures
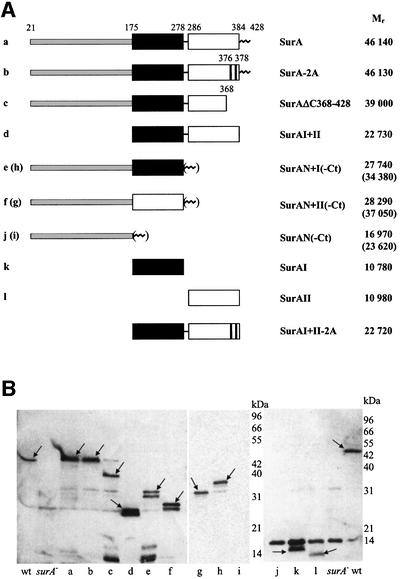
Fig. 1. Schematic representation (A) and periplasmic location (B) of SurA variants used in this study. (A) The mature proteins without signal peptide sequence (amino acids 1–20 in SurA) are shown and their relative predicted molecular weights (Mr) are given. Numbers refer to amino acid positions in the non-processed peptide sequence. Black box, parvulin-like inactive domain I; white box, parvulin-like active domain II. The N-terminal region of SurA is shaded light gray, the very C-terminus (Ct) of SurA (amino acids 385–428) is shown as a zig-zag line. The positions of alanine substitutions in SurA-2A and SurAI+II-2A are indicated. For all in vivo experiments, these fragments were preceded by the SurA signal peptide sequence so that they would be secreted into the periplasm (see Materials and methods). (B) Western blot detection of SurA variants in periplasmic fractions of surA– cells producing plasmid-encoded: (a) SurA; (b) SurA-2A; (c) SurAΔC368–428; (d) SurAI+II; (e) SurAN+I; (f) SurAN+II; (g) SurAN+II-Ct; (h) SurAN+I-Ct; (i) SurAN-Ct; (j) SurAN; (k) SurAI; and (l) SurAII. Periplasmic fractions of wild-type cells (wt) and surA– cells (surA–) served respectively as positive and negative controls. Extracts from equal amounts of cells were loaded in each lane, and western blot analysis, loading controls and controls for contamination by cytoplasmic fraction (data not shown) were performed as described in Materials and methods. Arrows indicate specific signals. A non-specific cross-reaction with lysozyme (14 kDa signal in the right panel) was observed occasionally in western blots with SurA antibodies. The wild-type sized signal in lane c was not observed in other experiments and results from a spill-over from the neighboring lane.
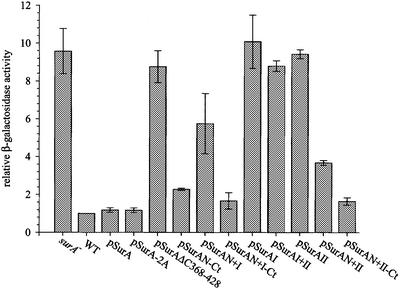
Fig. 2. EσE activity in surA– cells producing periplasmic SurA variants. EσE activity was assayed by monitoring β-galactosidase activity originating from a single chromosomal copy of the reporter fusion φλ[rpoHP3::lacZ]. The differential rates of β-galactosidase synthesis in surA– cells were normalized to the activity of wild-type cells. Results represent the average of at least two experiments.
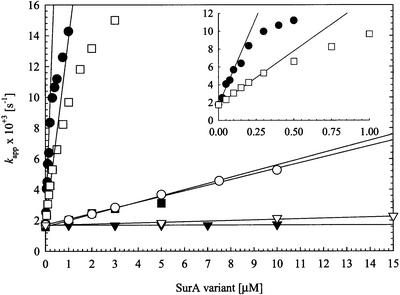
Fig. 3. Refolding of RCM-T1 by wild-type SurA (filled circles), the alanine double mutant SurA-2A (filled squares), the SurA fragments SurAI (domain I, filled triangles), SurAII (domain II, open circles), SurAI+II (domain I+II, open squares) and SurAI+II-2A (domain I+II-2A, open triangles) at 15°C. The dependence of the apparent rate of slow folding (Kapp) on the concentration of these SurA variants is shown. The concentration of unfolded RCM-T1 substrate was 0.5 µM for SurA and SurAI+II and 2.0 µM for SurA-2A and SurAI. The kcat/Km values (Table II) were obtained from the slopes of the indicated regression lines. The inset gives a more precise picture of the refolding of RCM-T1 by wild-type SurA and SurAI+II at concentrations between 0 and 1 µM. SurA proteins with an N-terminal region have the SurA signal sequence at their N-terminus and a C-terminal His6 tag; these proteins were purified from periplasmic fractions. All other fragments contained an N-terminal His6 tag and were purified from whole-cell extracts.
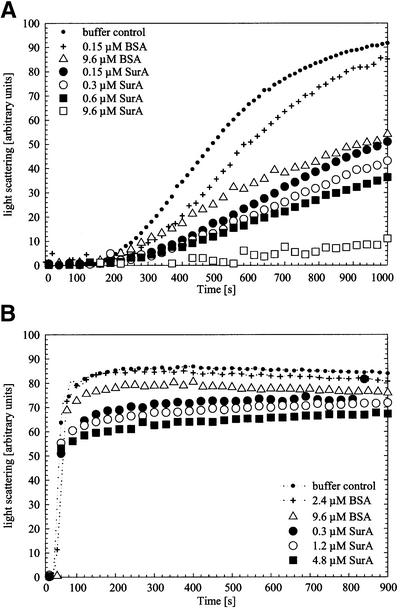
Fig. 4. Aggregation of citrate synthase during thermal stress (A) and during renaturation (B) in the absence of additional protein (buffer control) or in the presence of BSA or SurA in increasing concentrations. Light-scattering measurements of citrate synthase (0.15 µM monomer) were performed in 40 mM HEPES–KOH pH 7.5 at 43°C for thermal aggregation and in 50 mM Tris–HCl pH 8.0 at 25°C for renaturation experiments, respectively. To exclude effects that may be caused by the protein buffer (10 mM Tris–HCl pH 8.0, 300 mM NaCl, 5% glycerol), all samples were added to the assay in constant volume.
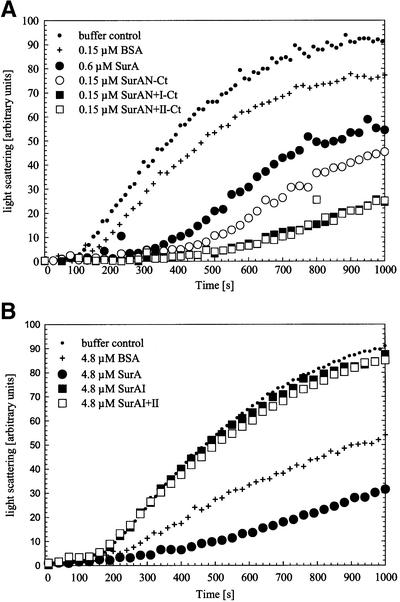
Fig. 5. Influence of SurA fragments with (A) and without (B) the N-terminal region of the protein on thermal aggregation of citrate synthase. Light-scattering measurements of citrate synthase (0.15 µM monomer) were performed without additional protein (buffer control) or in the presence of the indicated concentrations of BSA, SurA, SurAN-Ct, SurAN+I-Ct, SurAN+II-Ct, SurAI and SurAI+II. Buffer compositions and assay conditions were as in Figure 4.
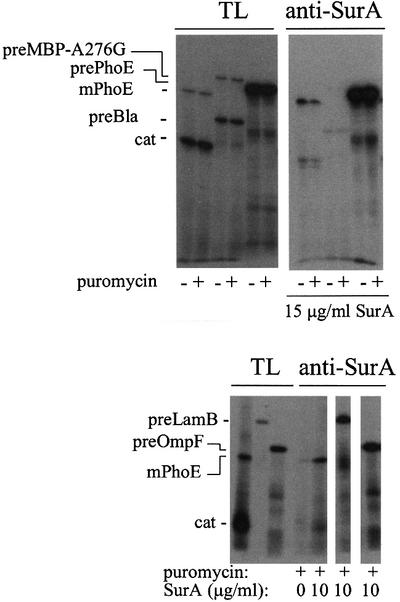
Fig. 6. Co-immunoprecipitation of in vitro synthezised proteins and SurA. Precursor and mature PhoE proteins (prePhoE and mPhoE), chloramphenicol acetyltransferase (cat) and the precursor proteins of LamB (preLamB), OmpF (preOmpF), the slow folding mutant maltose-binding protein (preMBP-A276G; Chun et al., 1993) and β-lactamase (preBla) were synthesized in vitro in the absence of SurA or with 10 or 15 µg/ml SurA added to the translation mixture and co-precipitated with anti-SurA antibodies as described in Materials and methods. The lanes with total translation products (TL) contain 10% of the total amount of translation mixtures used in the co-immunoprecipitations (anti-SurA). The amount of PhoE protein associating with SurA is similar in the absence or presence of puromycin (10 µM), which, where indicated, was added after protein synthesis to release the peptide chains from ribosomes.
References
-
- Bardwell J.C. and Beckwith,J. (1993) The bonds that tie: catalyzed disulfide bond formation. Cell, 74, 769–771. - PubMed
-
- Bardwell J.C., McGovern,K. and Beckwith,J. (1991) Identification of a protein required for disulfide bond formation in vivo. Cell, 67, 581–589. - PubMed
-
- Bosch D., Leunissen,J., Verbakel,J., de Jong,M., van Erp,H. and Tommassen,J. (1986) Periplasmic accumulation of truncated forms of outer-membrane PhoE protein of Escherichia coli K-12. J. Mol. Biol., 189, 449–455. - PubMed
-
- Bothmann H. and Plückthun,A. (1998) Selection for a periplasmic factor improving phage display and functional periplasmic expression. Nature Biotechnol., 16, 376–380. - PubMed
-
- Buchner J., Schmidt,M., Fuchs,M., Jaenicke,R., Rudolph,R., Schmid,F.X. and Kiefhaber,T. (1991) GroE facilitates refolding of citrate synthase by suppressing aggregation. Biochemistry, 30, 1586–1591. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
