

Presenilin-1 binds cytoplasmic epithelial cadherin, inhibits cadherin/p120 association, and regulates stability and function of the cadherin/catenin adhesion complex
- PMID: 11226248
- PMCID: PMC30147
- DOI: 10.1073/pnas.041603398
Presenilin-1 binds cytoplasmic epithelial cadherin, inhibits cadherin/p120 association, and regulates stability and function of the cadherin/catenin adhesion complex
Abstract
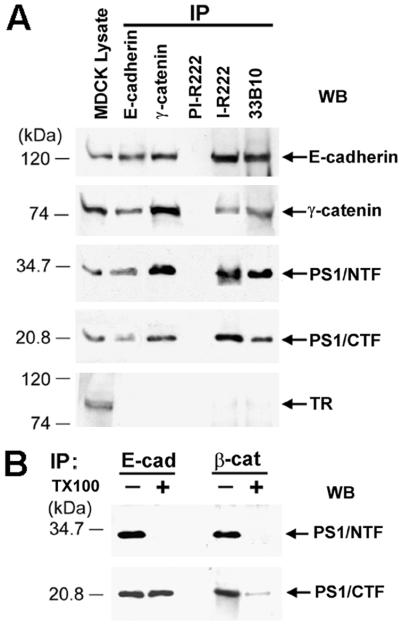
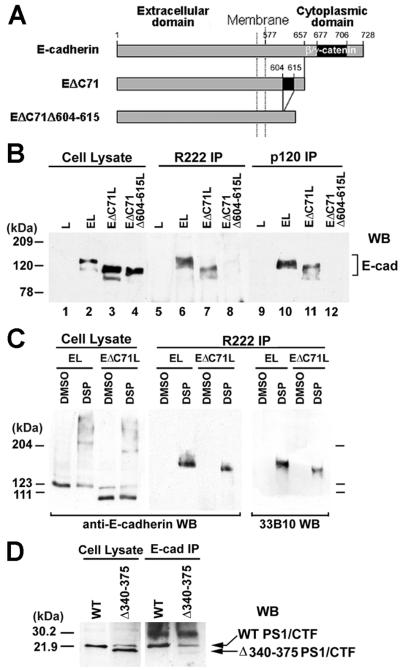
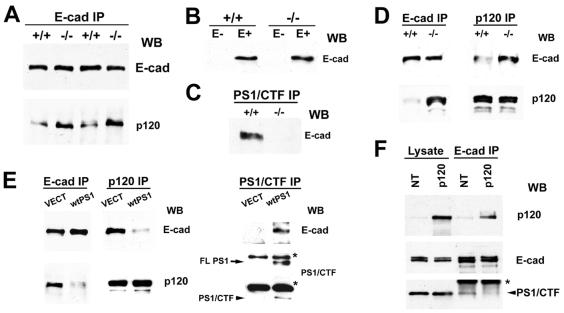
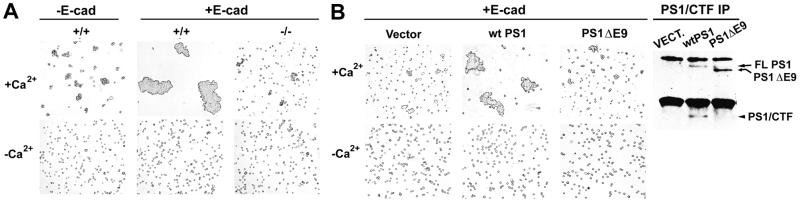
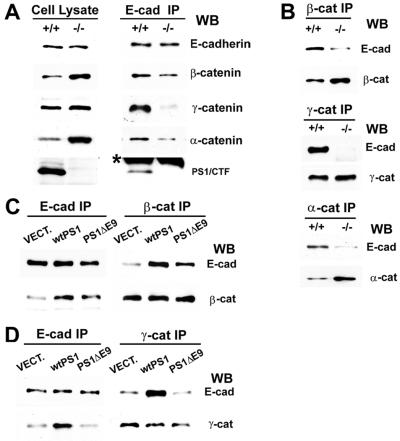
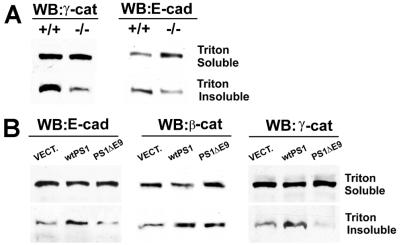
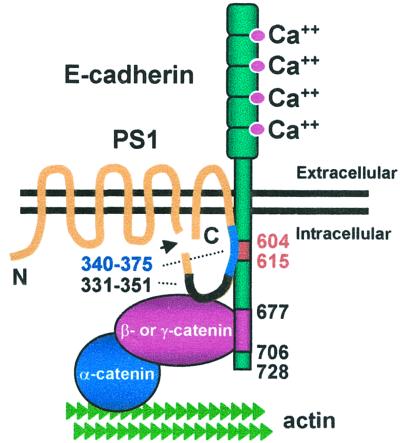
Here we show that presenilin-1 (PS1), a protein involved in Alzheimer's disease, binds directly to epithelial cadherin (E-cadherin). This binding is mediated by the large cytoplasmic loop of PS1 and requires the membrane-proximal cytoplasmic sequence 604-615 of mature E-cadherin. This sequence is also required for E-cadherin binding of protein p120, a known regulator of cadherin-mediated cell adhesion. Using wild-type and PS1 knockout cells, we found that increasing PS1 levels suppresses p120/E-cadherin binding, and increasing p120 levels suppresses PS1/E-cadherin binding. Thus PS1 and p120 bind to and mutually compete for cellular E-cadherin. Furthermore, PS1 stimulates E-cadherin binding to beta- and gamma-catenin, promotes cytoskeletal association of the cadherin/catenin complexes, and increases Ca(2+)-dependent cell-cell aggregation. Remarkably, PS1 familial Alzheimer disease mutant DeltaE9 increased neither the levels of cadherin/catenin complexes nor cell aggregation, suggesting that this familial Alzheimer disease mutation interferes with cadherin-based cell-cell adhesion. These data identify PS1 as an E-cadherin-binding protein and a regulator of E-cadherin function in vivo.
Figures
References
-
- Sherrington R, Rogaev E I, Liang Y, Rogaeva E A, Levesque G, Ikeda M, Chi H, Lin C, Li G, Holman K, et al. Nature (London) 1995;375:754–760. - PubMed
-
- Elder G A, Tezapsidis N, Carter J, Shioi J, Bouras C, Li H C, Johnston J M, Efthimiopoulos S, Friedrich V L, Jr, Robakis N K. J Neurosci Res. 1996;45:308–320. - PubMed
-
- Podlisny M B, Citron M, Amarante P, Sherrington R, Xia W, Zhang J, Diehl T, Levesque G, Fraser P, Haass C, et al. Neurobiol Dis. 1997;3:325–337. - PubMed
-
- Thinakaran G, Borchelt D R, Lee M K, Slunt H H, Spitzer L, Kim G, Ratovitsky T, Davenport F, Nordstedt C, Seeger M, et al. Neuron. 1996;17:181–190. - PubMed
-
- De Strooper B, Annaert W, Cupers P, Saftig P, Craessaerts K, Mumm J S, Schroeter E H, Schrijvers V, Wolfe M S, Ray W J, et al. Nature (London) 1999;398:518–522. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
