

Brain 5alpha-dihydroprogesterone and allopregnanolone synthesis in a mouse model of protracted social isolation
- PMID: 11226329
- PMCID: PMC30228
- DOI: 10.1073/pnas.051628598
Brain 5alpha-dihydroprogesterone and allopregnanolone synthesis in a mouse model of protracted social isolation
Abstract
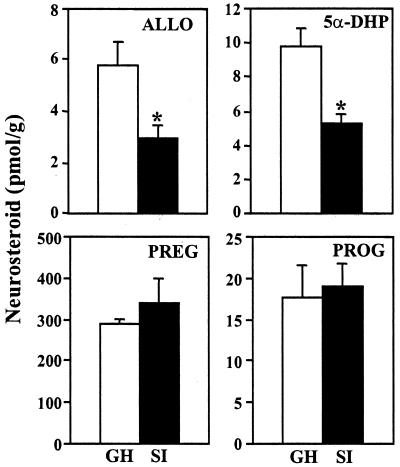
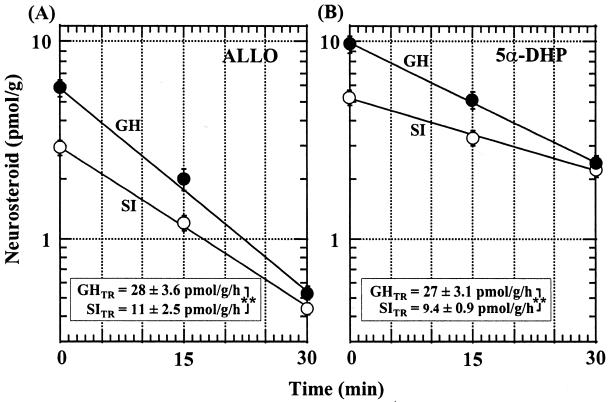
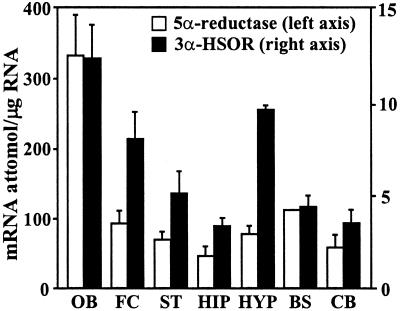
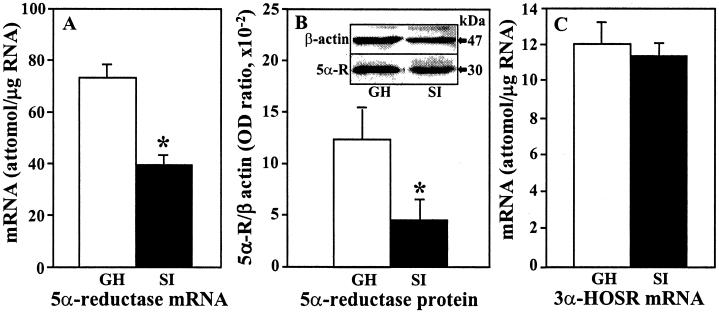
Allopregnanolone (ALLO), is a brain endogenous neurosteroid that binds with high affinity to gamma-aminobutyric acid type A (GABA(A)) receptors and positively modulates the action of GABA at these receptors. Unlike ALLO, 5alpha-dihydroprogesterone (5alpha-DHP) binds with high affinity to intracellular progesterone receptors that regulate DNA transcription. To investigate the physiological roles of ALLO and 5alpha-DHP synthesized in brain, we have adopted a mouse model involving protracted social isolation. In the frontal cortex of mice, socially isolated for 6 weeks, both neurosteroids were decreased by approximately 50%. After administration of (17beta)-17-(bis-1-methyl amino carbonyl) androstane-3,5-diene-3-carboxylic acid (SKF105,111), an inhibitor of the enzyme (5alpha-reductase Type I and II) that converts progesterone into 5alpha-DHP, the ALLO and 5alpha-DHP content of frontal cortex of both group-housed and socially isolated mice decreased exponentially to 10%-20% of control values in about 30 min. The fractional rate constants (k h(-1)) of ALLO and 5alpha-DHP decline multiplied by the ALLO and 5alpha-DHP concentrations at any given steady-state estimate the rate of synthesis required to maintain that steady state. After 6 weeks of social isolation, ALLO and 5alpha-DHP biosynthesis rates were decreased to 30% of the values calculated in group-housed mice. Moreover, in socially isolated mice, the expression of 5alpha-reductase Type I mRNA and protein was approximately 50% lower than in group-housed mice whereas 3alpha-hydroxysteroid oxidoreductase mRNA expression was equal in the two groups. Protracted social isolation in mice may provide a model to investigate whether 5alpha-DHP by a genomic action, and ALLO by a nongenomic mechanism down-regulate the action of drugs acting as agonists, partial agonists, or positive allosteric modulators of the benzodiazepine recognition sites expressed by GABA(A) receptors.
Figures
References
-
- Guidotti A, Costa E. Biol Psychiatry. 1998;44:856–873. - PubMed
-
- Compagnone N A, Mellon S H. Front Neuroendocrinol. 2000;21:1–56. - PubMed
-
- McEwen B S. Trends Pharmacol Sci. 1991;12:141–147. - PubMed
-
- Rupprecht R, Holsboer F. Trends Neurosci. 1999;22:410–416. - PubMed
-
- Matsumoto K, Uzunova V, Uzunov D P, Costa E, Guidotti A. Neuropharmacology. 1999;38:955–963. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
