

Reconstitution and functional comparison of purified GlpF and AqpZ, the glycerol and water channels from Escherichia coli
- PMID: 11226336
- PMCID: PMC30235
- DOI: 10.1073/pnas.051628098
Reconstitution and functional comparison of purified GlpF and AqpZ, the glycerol and water channels from Escherichia coli
Abstract
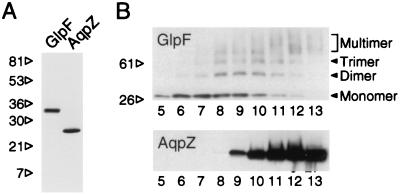
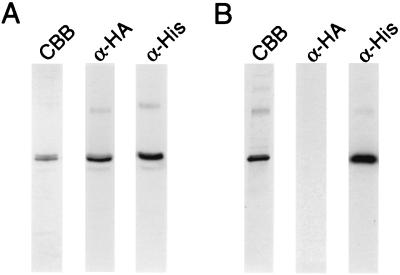
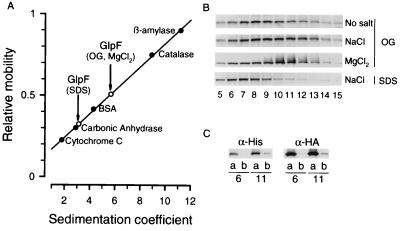
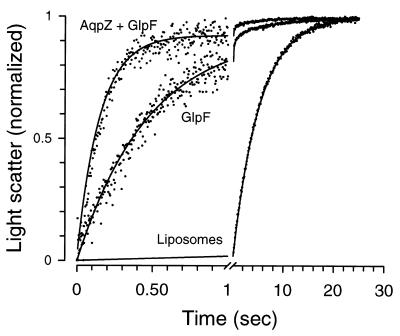
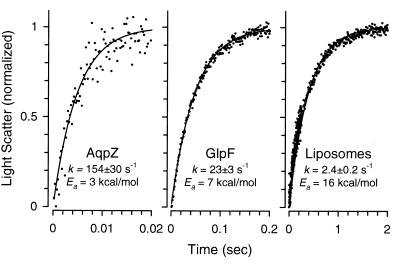
A large family of membrane channel proteins selective for transport of water (aquaporins) or water plus glycerol (aquaglyceroporins) has been found in diverse life forms. Escherichia coli has two members of this family-a water channel, AqpZ, and a glycerol facilitator, GlpF. Despite having similar primary amino acid sequences and predicted structures, the oligomeric state and solute selectivity of AqpZ and GlpF are disputed. Here we report biochemical and functional characterizations of affinity-purified GlpF and compare it to AqpZ. Histidine-tagged (His-GlpF) and hemagglutinin-tagged (HA-GlpF) polypeptides encoded by a bicistronic construct were expressed in bacteria. HA-GlpF and His-GlpF appear to form oligomers during Ni-nitrilotriacetate affinity purification. Sucrose gradient sedimentation analyses showed that the oligomeric state of octyl glucoside-solubilized GlpF varies: low ionic strength favors subunit dissociation, whereas Mg(2+) stabilizes tetrameric assembly. Reconstitution of affinity-purified GlpF into proteoliposomes increases glycerol permeability more than 100-fold and water permeability up to 10-fold compared with control liposomes. Glycerol and water permeability of GlpF both occur with low Arrhenius activation energies and are reversibly inhibited by HgCl(2). Our studies demonstrate that, unlike AqpZ, a water-selective stable tetramer, purified GlpF exists in multiple oligomeric forms under nondenaturing conditions and is highly permeable to glycerol but less well permeated by water.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
