

The mouse anterior chamber angle and trabecular meshwork develop without cell death
- PMID: 11228591
- PMCID: PMC31337
- DOI: 10.1186/1471-213x-1-3
The mouse anterior chamber angle and trabecular meshwork develop without cell death
Abstract
Background: The iridocorneal angle forms in the mammalian eye from undifferentiated mesenchyme between the root of the iris and cornea. A major component is the trabecular meshwork, consisting of extracellular matrix organized into a network of beams, covered in trabecular endothelial cells. Between the beams, channels lead to Schlemm's canal for the drainage of aqueous humor from the eye into the blood stream. Abnormal development of the iridocorneal angle that interferes with ocular fluid drainage can lead to glaucoma in humans. Little is known about the precise mechanisms underlying angle development. There are two main hypotheses. The first proposes that morphogenesis involves mainly cell differentiation, matrix deposition and assembly of the originally continuous mesenchymal mass into beams, channels and Schlemm's canal. The second, based primarily on rat studies, proposes that cell death and macrophages play an important role in forming channels and beams. Mice provide a potentially useful model to understand the origin and development of angle structures and how defective development leads to glaucoma. Few studies have assessed the normal structure and development of the mouse angle. We used light and electron microscopy and a cell death assay to define the sequence of events underlying formation of the angle structures in mice.
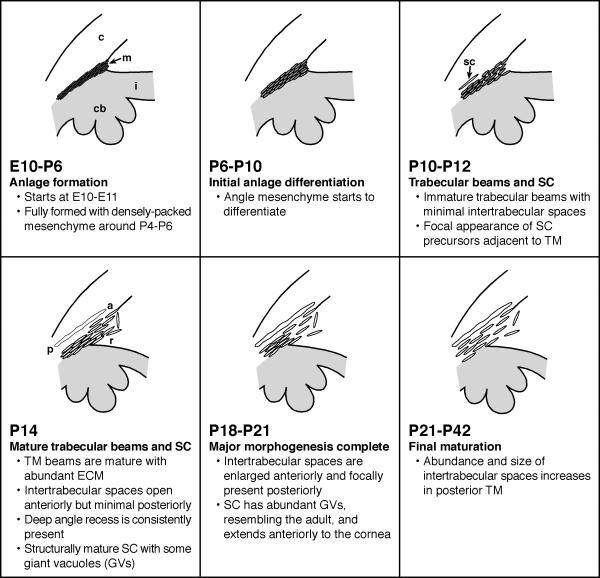
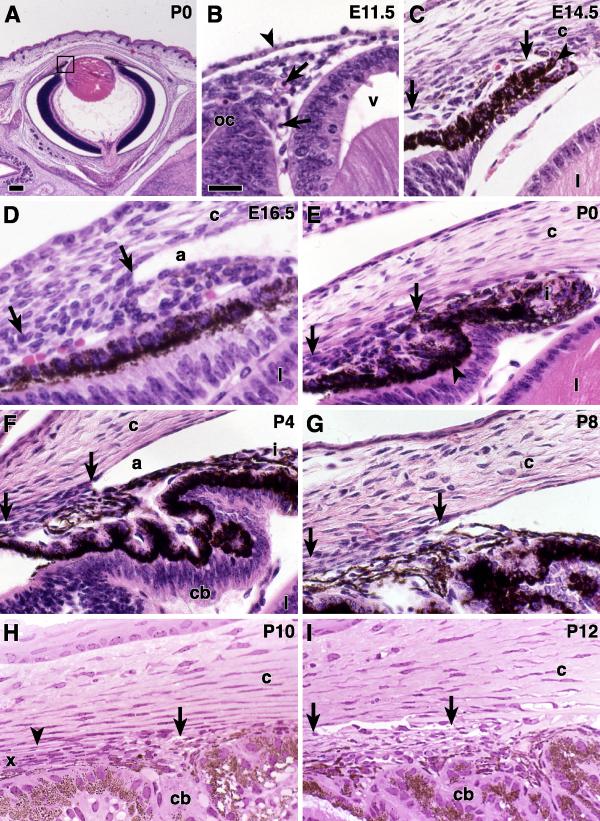
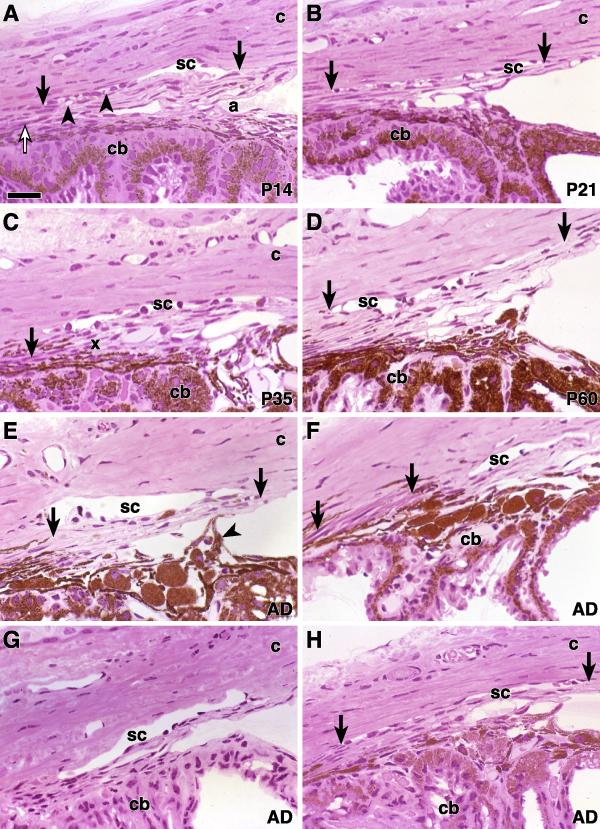
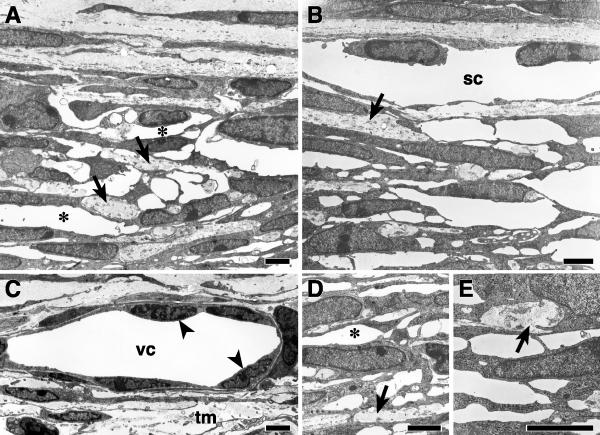
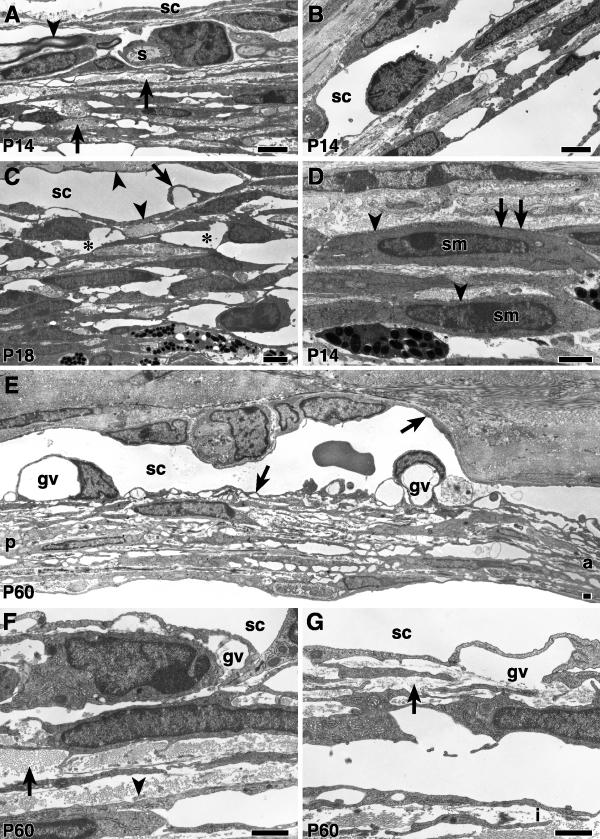
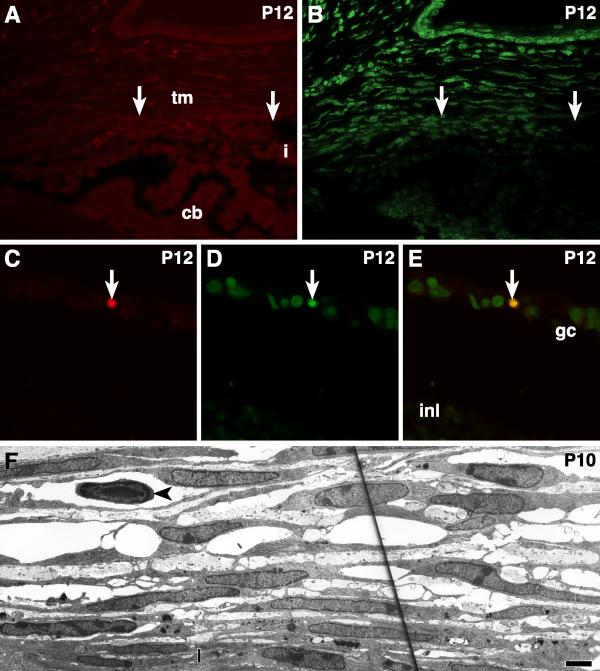
Results: The mouse angle structures and developmental sequence are similar to those in humans. Cell death was not detectable during the period of trabecular channel and beam formation.
Conclusions: These results support morphogenic mechanisms involving organization of cellular and extracellular matrix components without cell death or atrophy.
Figures
References
-
- Ritch R, Shields MB, Krupin T. The Glaucomas, Clinical Science, 2nd edn. St Louis, MO: Mosby-Year Book; 1996.
-
- Caprioli J. The ciliary epithelia and aqueous humor. Adler's Physiology of the Eye Edited by William M. Hart J, 9th ed. pp. 228-247. St. Louis: Mosby Year Book; 1992. pp. 228–247.
-
- Hart WM. Intraocular Pressure. Adler's Physiology of the Eye Edited by William M. Hart J, 9th ed. pp. 248-267. St. Louis: Mosby Year Book; 1992. pp. 248–267.
-
- Bill A. Uveoscleral drainage of aqueous humor: physiology and pharmacology. Prog Clin Biol Res. 1989;312:417–427. - PubMed
-
- Mann IC. The Development of the Human Eye, First edn. Cambridge: Cambridge University Press; 1928.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
