

Recruitment, activation and retention of caspases-9 and -3 by Apaf-1 apoptosome and associated XIAP complexes
- PMID: 11230124
- PMCID: PMC145489
- DOI: 10.1093/emboj/20.5.998
Recruitment, activation and retention of caspases-9 and -3 by Apaf-1 apoptosome and associated XIAP complexes
Abstract
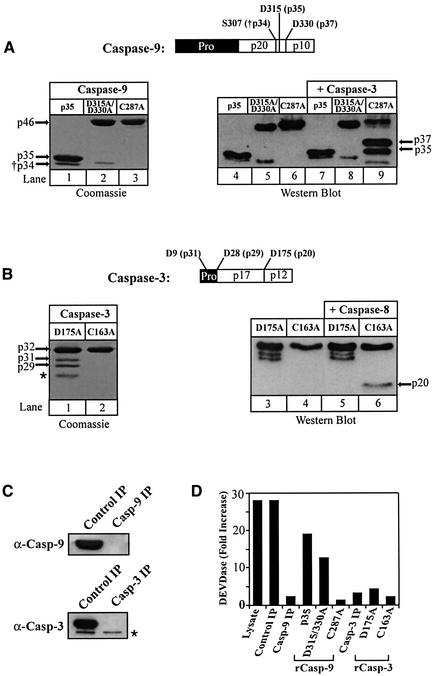
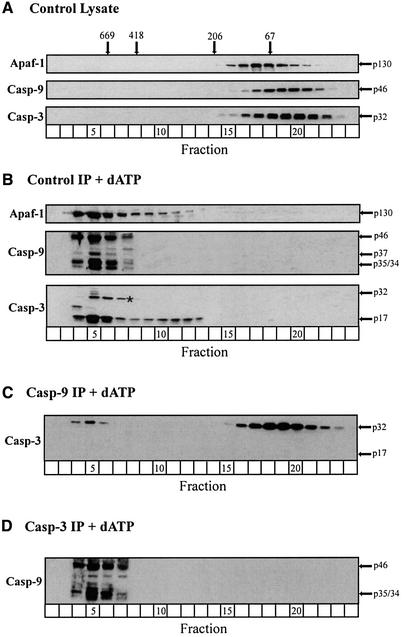
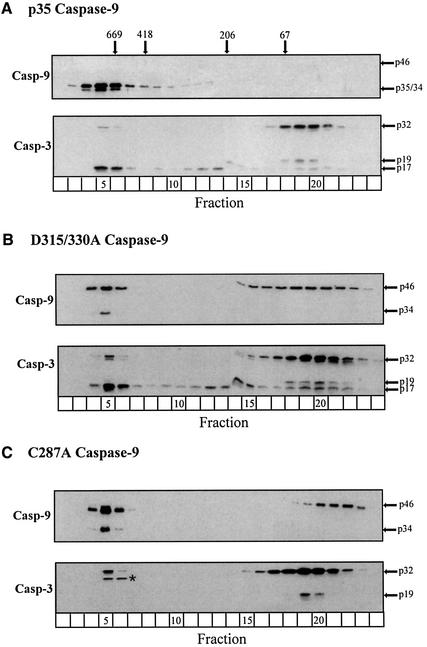
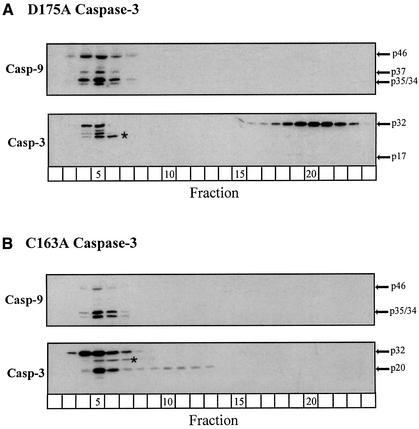
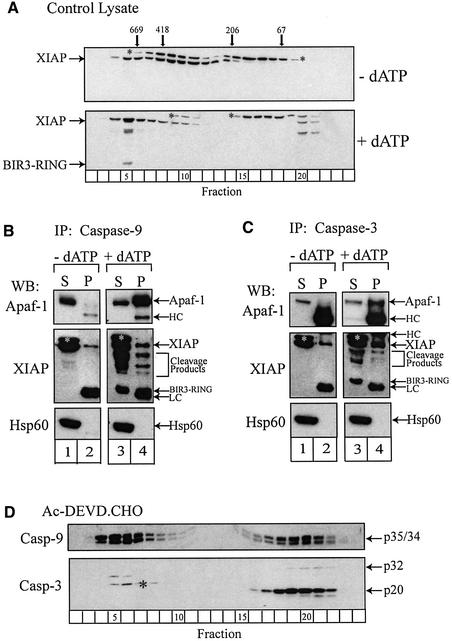
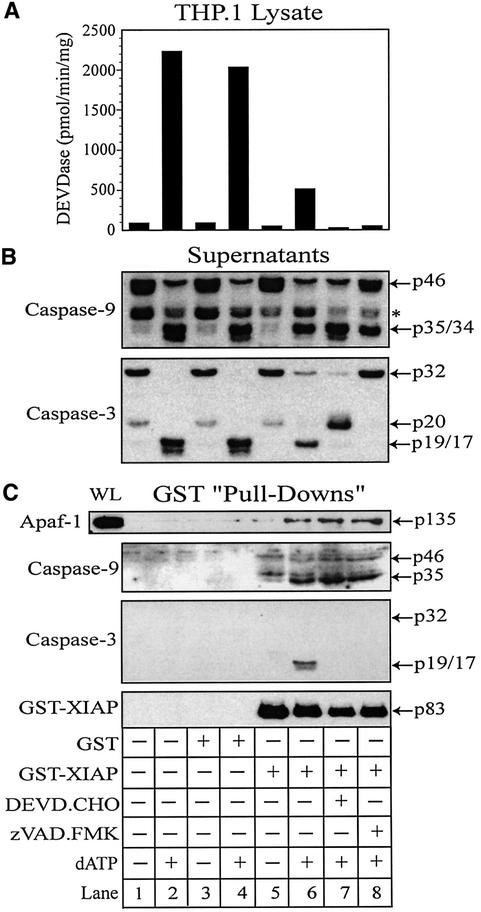
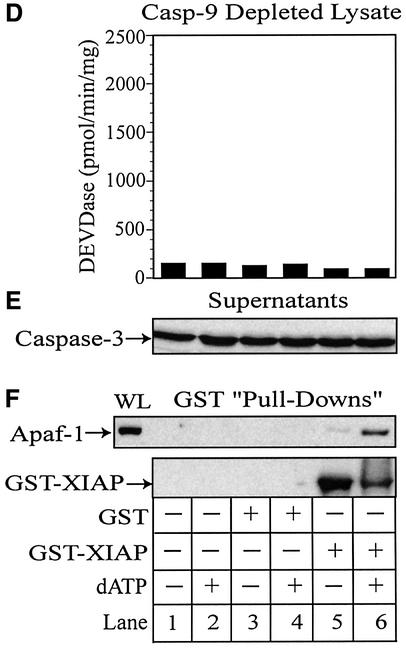
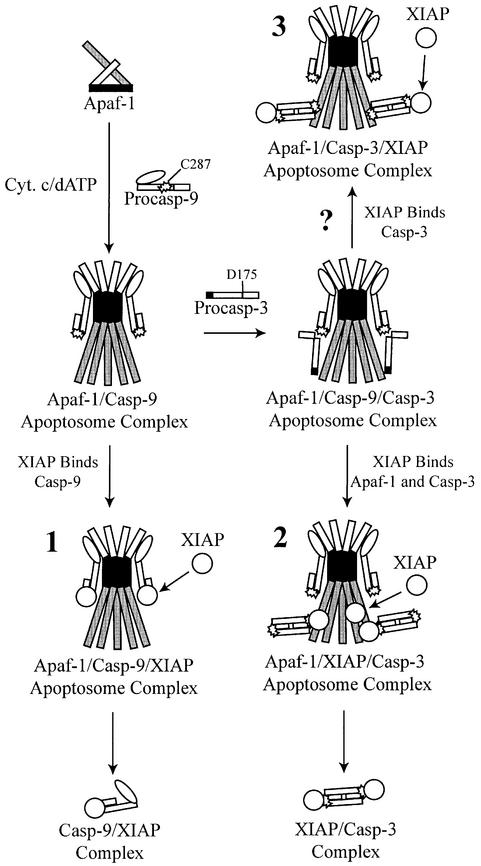
During apoptosis, release of cytochrome c initiates dATP-dependent oligomerization of Apaf-1 and formation of the apoptosome. In a cell-free system, we have addressed the order in which apical and effector caspases, caspases-9 and -3, respectively, are recruited to, activated and retained within the apoptosome. We propose a multi-step process, whereby catalytically active processed or unprocessed caspase-9 initially binds the Apaf-1 apoptosome in cytochrome c/dATP-activated lysates and consequently recruits caspase-3 via an interaction between the active site cysteine (C287) in caspase-9 and a critical aspartate (D175) in caspase-3. We demonstrate that XIAP, an inhibitor-of-apoptosis protein, is normally present in high molecular weight complexes in unactivated cell lysates, but directly interacts with the apoptosome in cytochrome c/dATP-activated lysates. XIAP associates with oligomerized Apaf-1 and/or processed caspase-9 and influences the activation of caspase-3, but also binds activated caspase-3 produced within the apoptosome and sequesters it within the complex. Thus, XIAP may regulate cell death by inhibiting the activation of caspase-3 within the apoptosome and by preventing release of active caspase-3 from the complex.
Figures
References
-
- Bratton S.B., MacFarlane,M., Cain,K. and Cohen,G.M. (2000) Protein complexes activate distinct caspase cascades in death receptor and stress-induced apoptosis. Exp. Cell Res., 256, 27–33. - PubMed
-
- Cain K., Brown,D.G., Langlais,C. and Cohen,G.M. (1999) Caspase activation involves the formation of the aposome, a large (∼700 kDa) caspase-activating complex. J. Biol. Chem., 274, 22686–22692. - PubMed
-
- Cain K., Bratton,S.B., Langlais,C., Walker,G., Brown,D.G., Sun,X.M. and Cohen,G.M. (2000) Apaf-1 oligomerizes into biologically active ∼700 kDa and inactive ∼1.4 MDa apoptosome complexes. J. Biol. Chem., 275, 6067–6070. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
