

Arabidopsis glucosidase I mutants reveal a critical role of N-glycan trimming in seed development
- PMID: 11230125
- PMCID: PMC145462
- DOI: 10.1093/emboj/20.5.1010
Arabidopsis glucosidase I mutants reveal a critical role of N-glycan trimming in seed development
Abstract
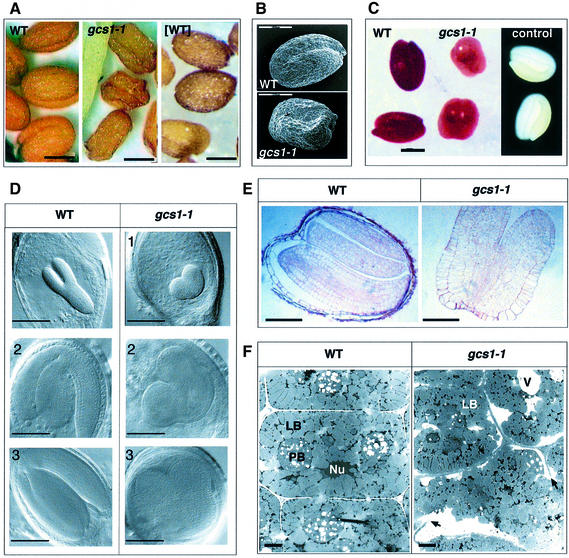
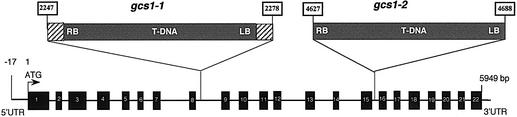
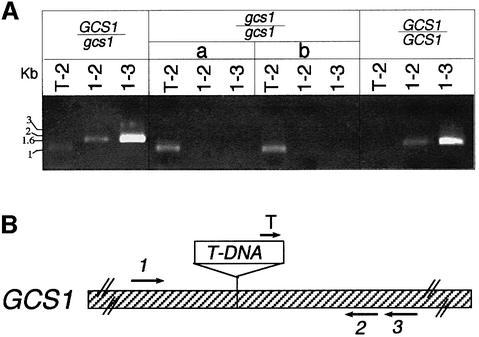
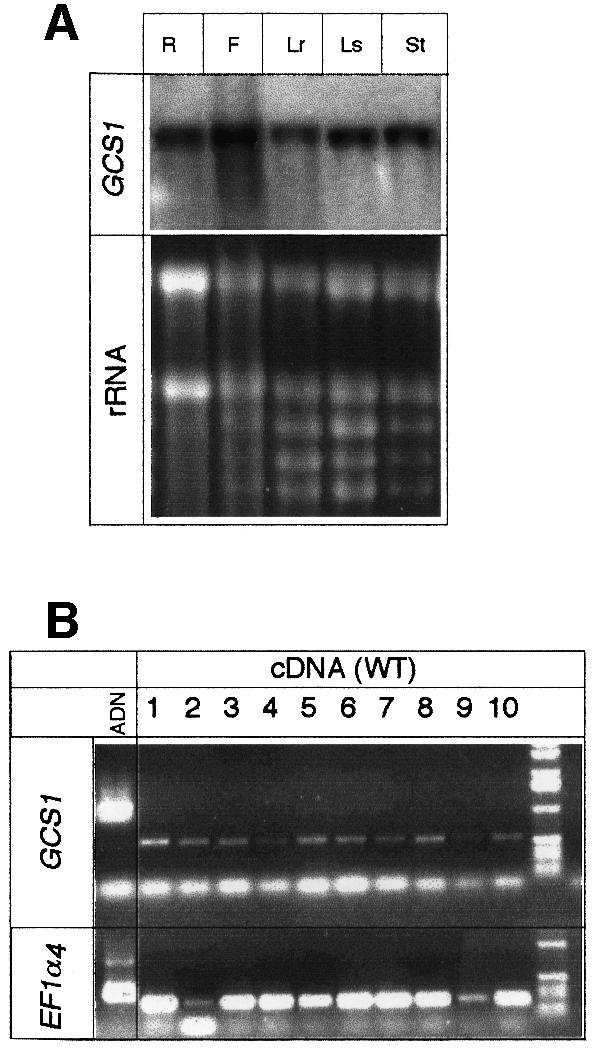
Glycoproteins with asparagine-linked (N-linked) glycans occur in all eukaryotic cells. The function of their glycan moieties is one of the central problems in contemporary cell biology. N-glycosylation may modify physicochemical and biological protein properties such as conformation, degradation, intracellular sorting or secretion. We have isolated and characterized two allelic Arabidopsis mutants, gcs1-1 and gcs1-2, which produce abnormal shrunken seeds, blocked at the heart stage of development. The mutant seeds accumulate a low level of storage proteins, have no typical protein bodies, display abnormal cell enlargement and show occasional cell wall disruptions. The mutated gene has been cloned by T-DNA tagging. It codes for a protein homologous to animal and yeast alpha-glucosidase I, an enzyme that controls the first committed step for N-glycan trimming. Biochemical analyses have confirmed that trimming of the alpha1,2- linked glucosyl residue constitutive of the N-glycan precursor is blocked in this mutant. These results demonstrate the importance of N-glycan trimming for the accumulation of seed storage proteins, the formation of protein bodies, cell differentiation and embryo development.
Figures

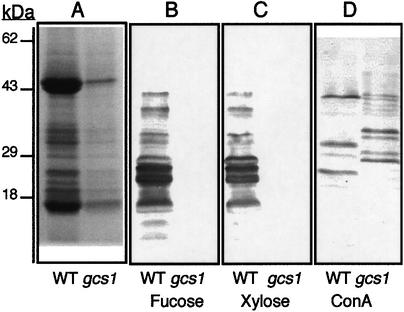
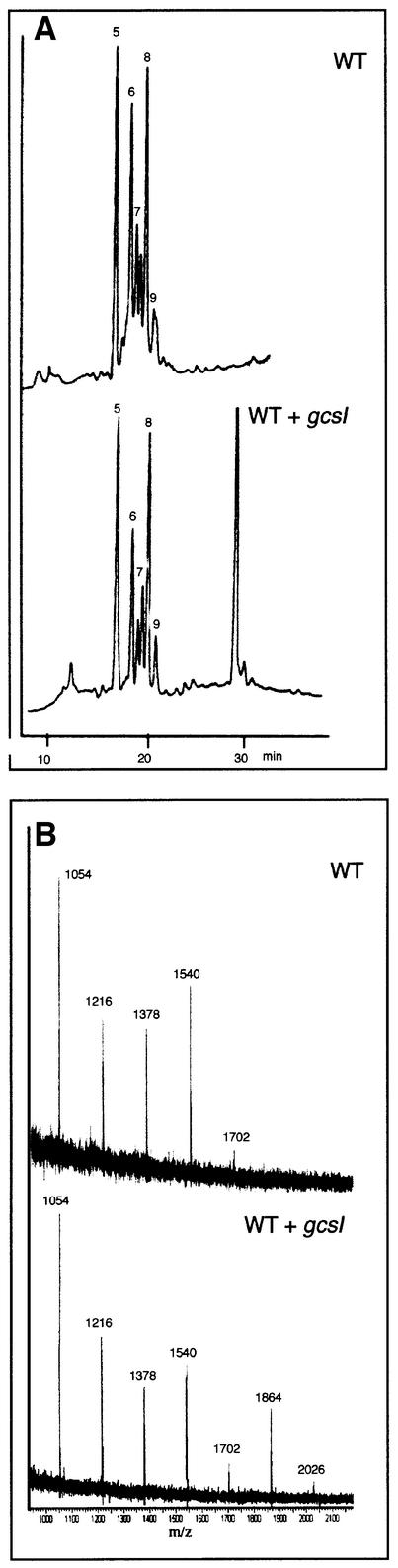
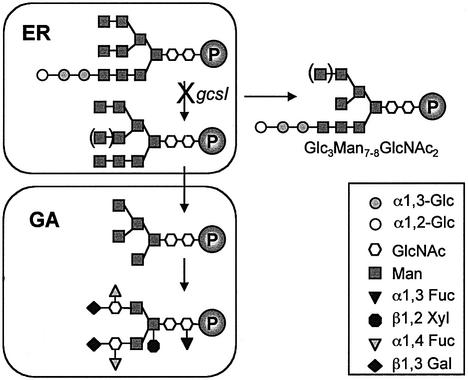
References
-
- Apweiler R., Hermjakob,H. and Sharon,N. (1999) On the frequency of protein glycosylation, as deduced from analysis of the SWISS-PROT database. Biochim. Biophys. Acta, 1473, 4–8. - PubMed
-
- Bardor M., Faye,L. and Lerouge,P. (1999) Analysis of the N-glycosylation of recombinant glycoproteins produced in transgenic plants. Trends Plant Sci., 4, 376–380. - PubMed
-
- Baus A.D., Franzmann,L. and Meinke,D.W. (1986) Growth of arrested embryos from lethal mutants of Arabidopsis thaliana. Theor. Appl. Genet., 72, 577–586. - PubMed
-
- Bechtold N., Ellis,J. and Pelletier,G. (1993) In planta Agrobacterium mediated gene transfer by infiltration of adult Arabidopsis thaliana plants. C. R. Acad. Sci. Paris, 316, 1194–1199.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
