

The NAF domain defines a novel protein-protein interaction module conserved in Ca2+-regulated kinases
- PMID: 11230129
- PMCID: PMC145464
- DOI: 10.1093/emboj/20.5.1051
The NAF domain defines a novel protein-protein interaction module conserved in Ca2+-regulated kinases
Abstract
The Arabidopsis calcineurin B-like calcium sensor proteins (AtCBLs) interact with a group of serine-threonine protein kinases (AtCIPKs) in a calcium-dependent manner. Here we identify a 24 amino acid domain (NAF domain) unique to these kinases as being required and sufficient for interaction with all known AtCBLs. Mutation of conserved residues either abolished or significantly diminished the affinity of AtCIPK1 for AtCBL2. Comprehensive two-hybrid screens with various AtCBLs identified 15 CIPKs as potential targets of CBL proteins. Database analyses revealed additional kinases from Arabidopsis and other plant species harbouring the NAF interaction module. Several of these kinases have been implicated in various signalling pathways mediating responses to stress, hormones and environmental cues. Full-length CIPKs show preferential interaction with distinct CBLs in yeast and in vitro assays. Our findings suggest differential interaction affinity as one of the mechanisms generating the temporal and spatial specificity of calcium signals within plant cells and that different combinations of CBL-CIPK proteins contribute to the complex network that connects various extracellular signals to defined cellular responses.
Figures






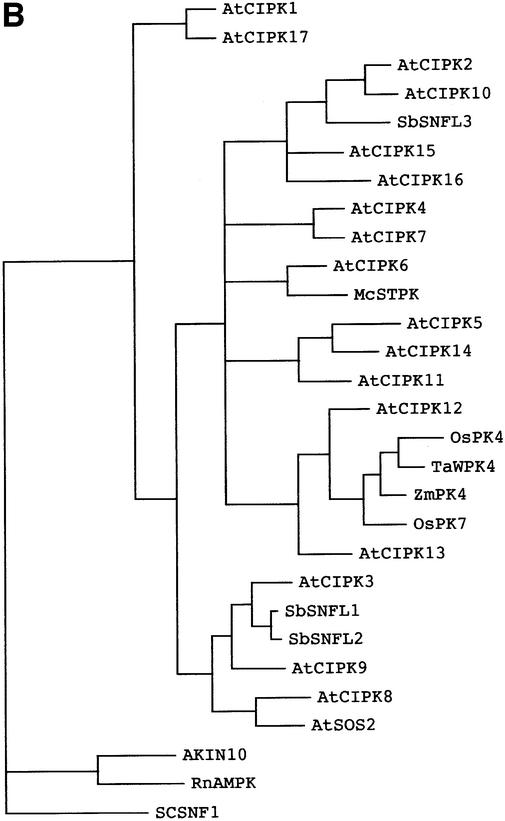
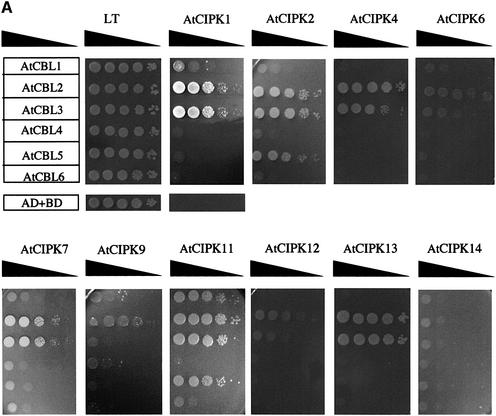

References
-
- Annen F. and Stockhaus,J. (1998) Characterisation of a Sorghum bicolor gene family encoding putative protein kinases with a high similarity to the yeast SNF1 protein kinase. Plant Mol. Biol., 36, 529–539. - PubMed
-
- Bootman M.D., Berridge,M.J. and Lipp,P. (1997) Cooking with calcium: the recipes for composing global signals from elementary events. Cell, 91, 367–373. - PubMed
-
- Chen C.-K., Inglese,J., Lefkowitz,R.J. and Hurley,J.B. (1995) Ca2+-dependent interaction of recoverin with rhodopsin kinase. J. Biol. Chem., 270, 18060–18066. - PubMed
-
- Clapham D.E. (1995) Calcium signalling. Cell, 80, 259–268. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
