

Regulation of the Sko1 transcriptional repressor by the Hog1 MAP kinase in response to osmotic stress
- PMID: 11230135
- PMCID: PMC145460
- DOI: 10.1093/emboj/20.5.1123
Regulation of the Sko1 transcriptional repressor by the Hog1 MAP kinase in response to osmotic stress
Abstract
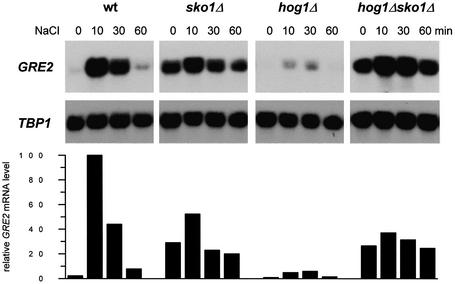
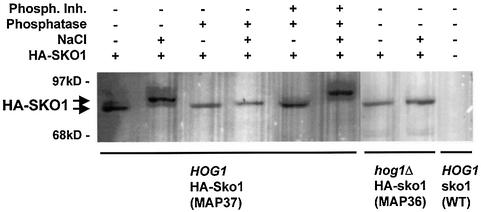
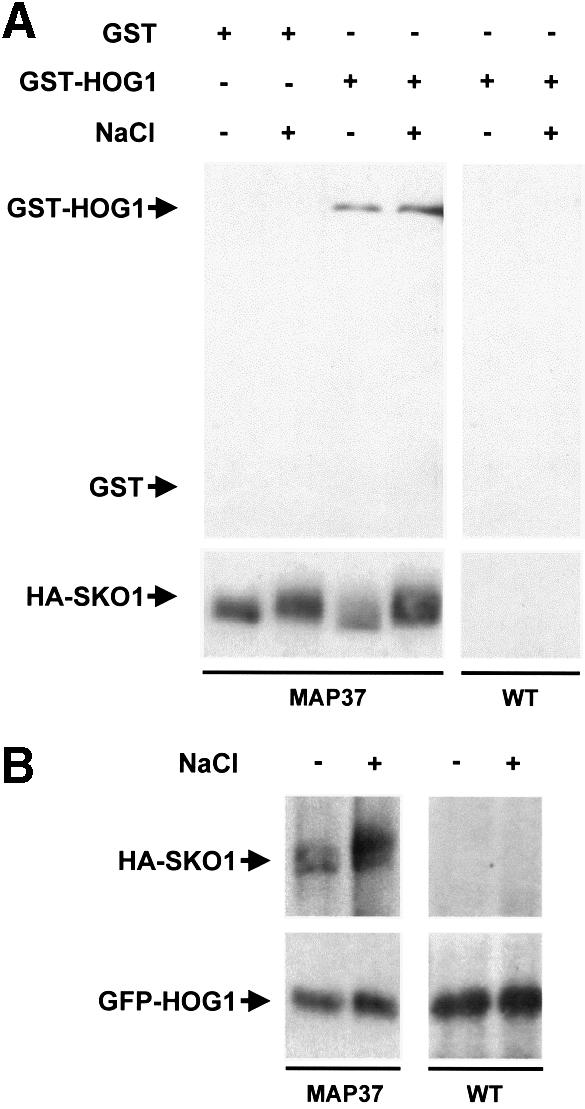
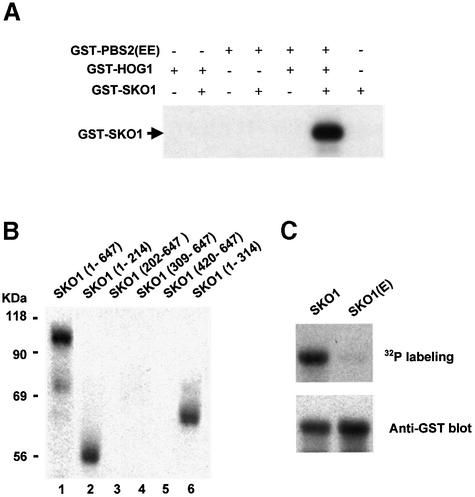
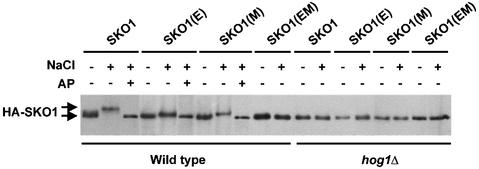
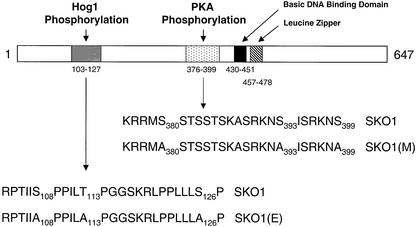
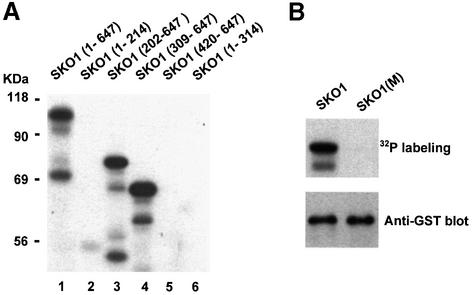
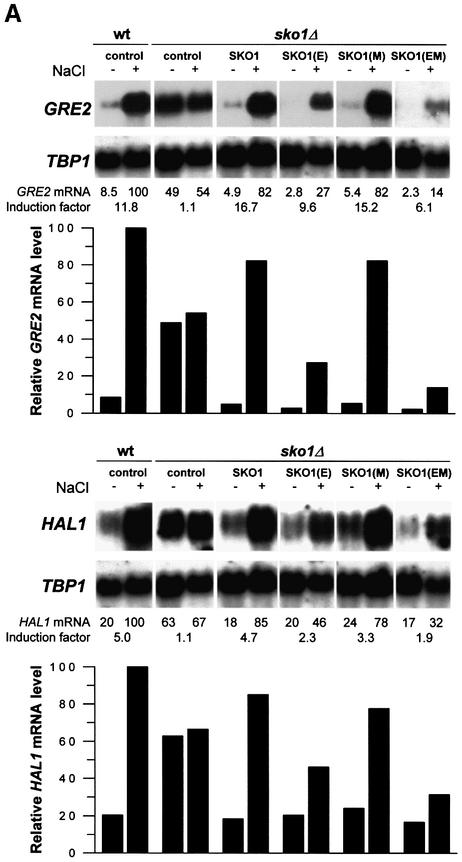
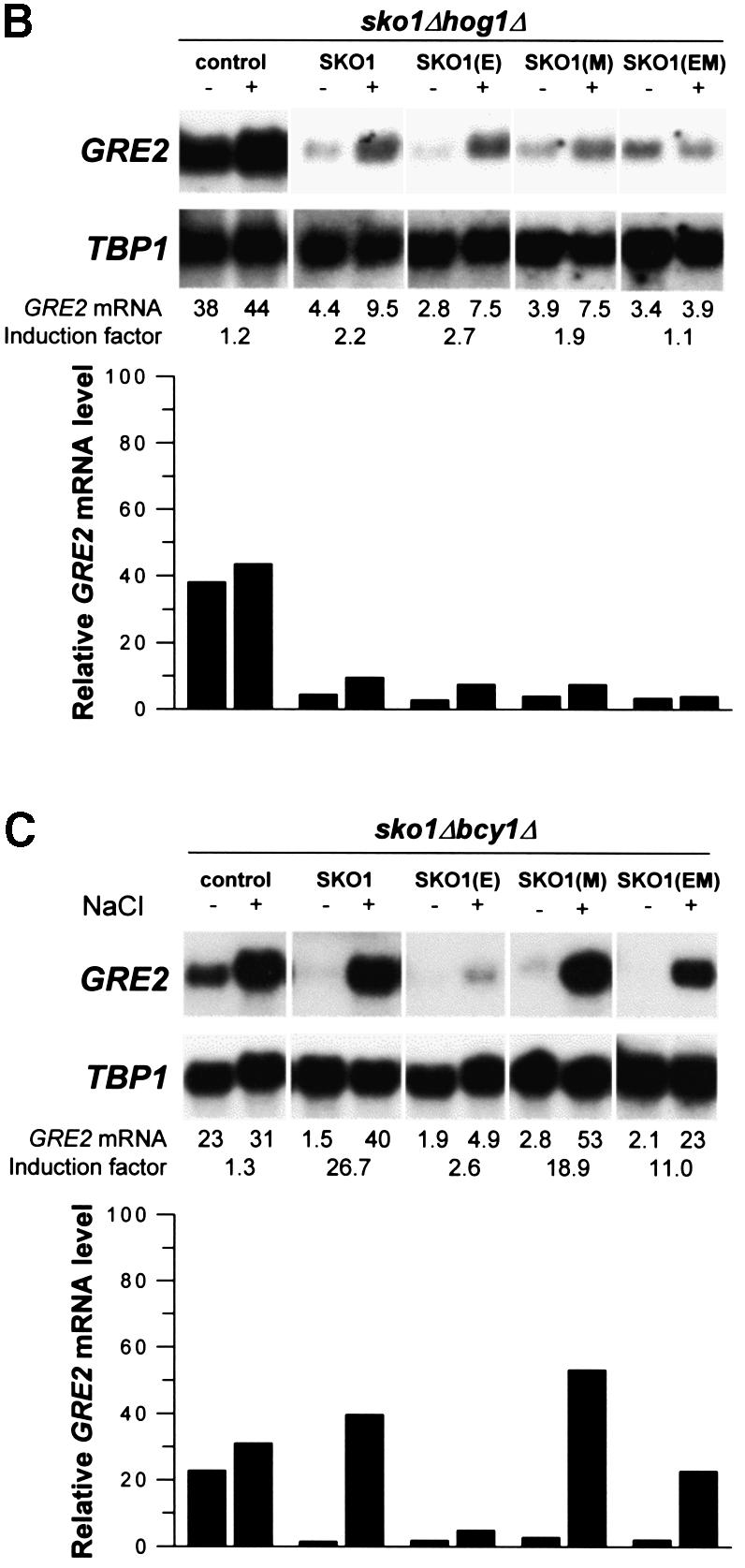
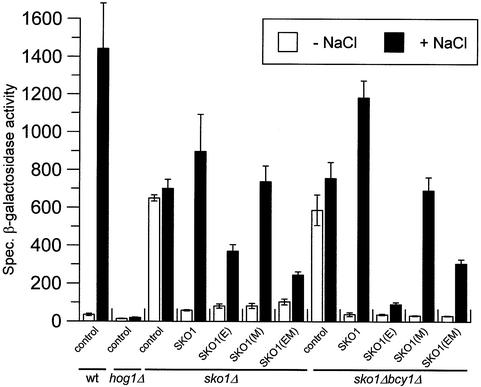
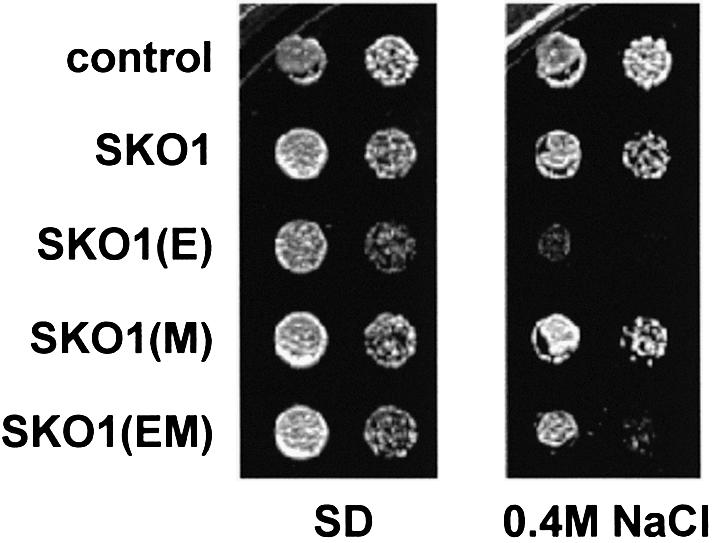
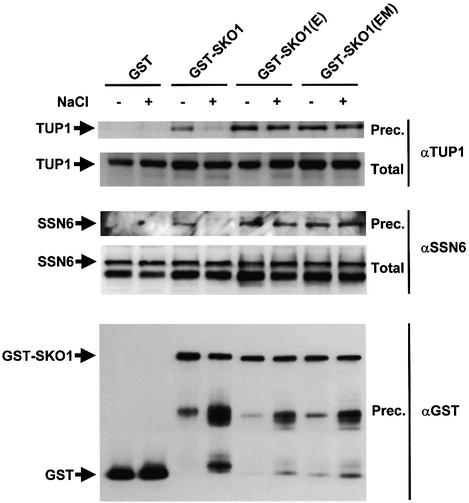
Exposure of yeast to increases in extracellular osmolarity activates the Hog1 mitogen-activated protein kinase (MAPK), which is essential for the induction of gene expression required for cell survival upon osmotic stress. Several genes are regulated in response to osmotic stress by Sko1, a transcriptional repressor of the ATF/CREB family. We show by in vivo coprecipitation and phosphorylation studies that Sko1 and Hog1 interact and that Sko1 is phosphorylated upon osmotic stress in a Hog1-dependent manner. Hog1 phosphorylates Sko1 in vitro at multiple sites within the N-terminal region. Phosphorylation of Sko1 disrupts the Sko1-Ssn6-Tup1 repressor complex, and consistently, a mutant allele of Sko1, unphosphorylatable by Hog1, exhibits less derepression than the wild type. Interestingly, Sko1 repressor activity is further enhanced in strains with high protein kinase A (PKA) activity. PKA phosphorylates Sko1 near the bZIP domain and mutation of these sites eliminates modulation of Sko1 responses to high PKA activity. Thus, Sko1 transcriptional repression is controlled directly by the Hog1 MAPK in response to stress, and this effect is further modulated by an independent signaling mechanism through the PKA pathway.
Figures
References
-
- Boguslawski G. (1992) PBS2, a yeast gene encoding a putative protein kinase, interacts with the RAS2 pathway and affects osmotic sensitivity of Saccharomyces cerevisiae. J. Gen. Microbiol., 138, 2425–2432. - PubMed
-
- Brewster J.L., de Valoir,T., Dwyer,N.D., Winter,E. and Gustin,M.C. (1993) An osmosensing signal transduction pathway in yeast. Science, 259, 1760–1763. - PubMed
-
- Carlson M. and Botstein,D. (1982) Two differentially regulated mRNAs with different 5′-ends encode secreted and intracellular forms of yeast invertase. Cell, 28, 145–154. - PubMed
-
- DeVit M.J. and Johnston,M. (1999) The nuclear exportin Msn5 is required for nuclear export of the Mig1 glucose repressor of Saccharomyces cerevisiae. Curr. Biol., 9, 1231–1241. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
