

Activation of the Notch-regulated transcription factor CBF1/RBP-Jkappa through the 13SE1A oncoprotein
- PMID: 11230145
- PMCID: PMC312632
- DOI: 10.1101/gad.189301
Activation of the Notch-regulated transcription factor CBF1/RBP-Jkappa through the 13SE1A oncoprotein
Abstract
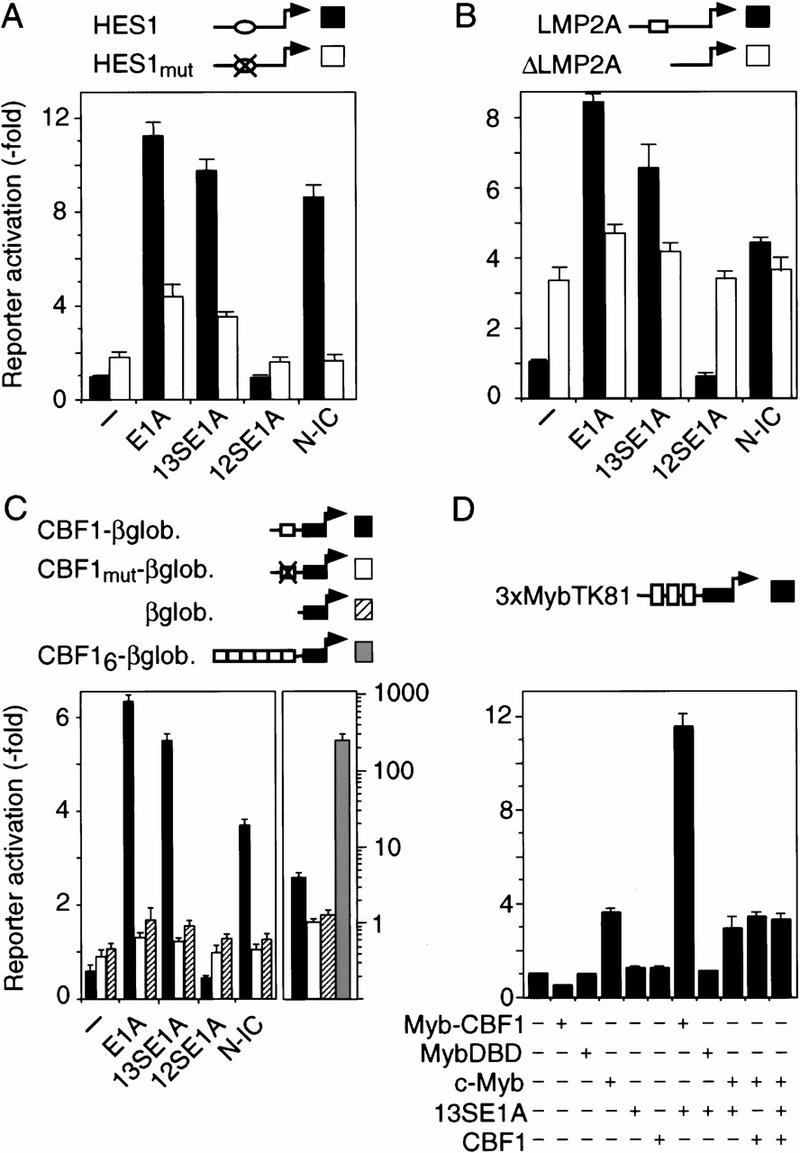
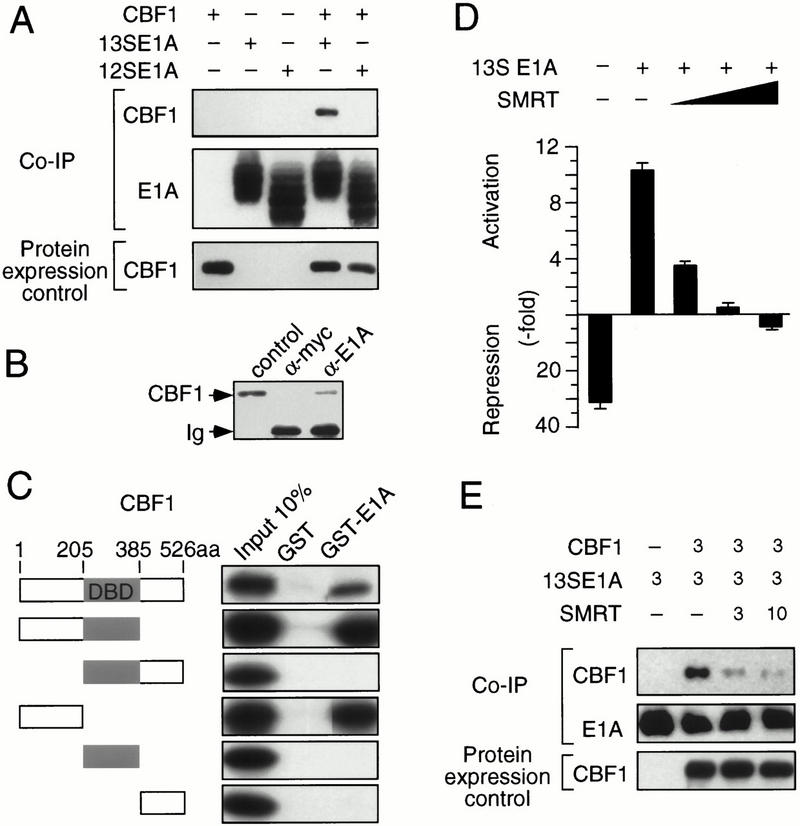
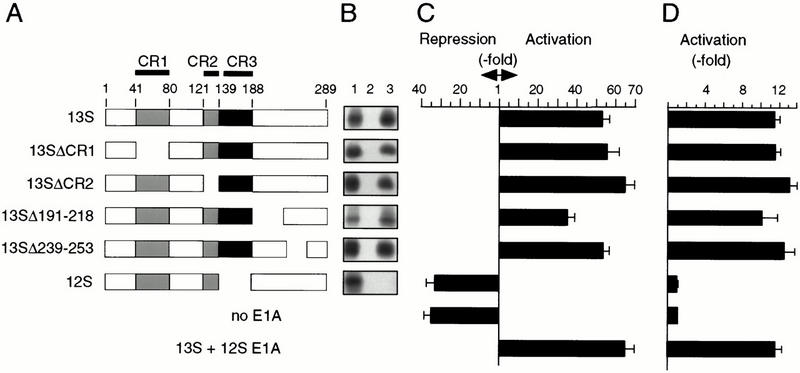
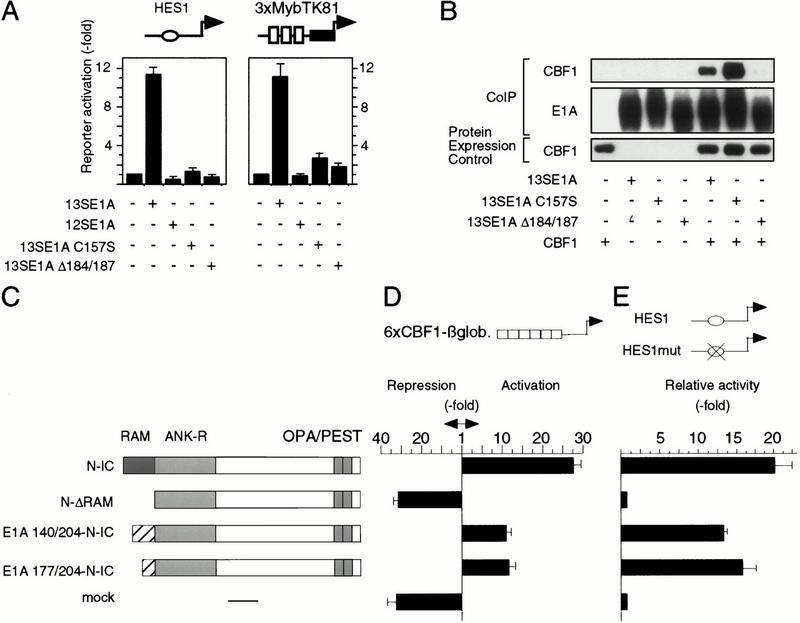
Signaling through the Notch pathway controls cell growth and differentiation in metazoans. Following binding of its ligands, the intracellular part of the cell surface Notch1 receptor (Notch1-IC) is released and translocates to the nucleus, where it alters the function of the DNA-binding transcription factor CBF1/RBP-Jkappa. As a result, CBF1/RBP-Jkappa is converted from a repressor to an activator of gene transcription. Similarly, the Epstein Barr viral oncoprotein EBNA2, which is required for B-cell immortalization, activates genes through CBF1. Moreover, the TAN-1 and int-3 oncogenes represent activated versions of Notch1 and Notch4, respectively. Here, we show that the adenoviral oncoprotein 13S E1A also binds to CBF1/RBP-Jkappa, displaces associated corepressor complexes, and activates CBF1/RBP-Jkappa-dependent gene expression. Our results suggest that the central role of the Notch-CBF1/RBP-Jkappa signaling pathway in cell fate decisions renders it susceptible to pathways of viral replication and oncogenic conversion.
Figures
References
-
- Artavanis-Tsakonas S, Matsuno K, Fortini ME. Notch signaling. Science. 1995;268:225–232. - PubMed
-
- Artavanis-Tsakonas S, Rand MD, Lake RJ. Notch signaling: Cell fate control and signal integration in development. Science. 1999;284:770–776. - PubMed
-
- Boyer TG, Martin ME, Lees E, Ricciardi RP, Berk AJ. Mammalian Srb/mediator complex is targeted by adenovirus E1A protein. Nature. 1999;399:276–279. - PubMed
-
- Brou C, Logeat F, Lecourtois M, Vandekerckhove J, Kourilsky P, Schweisguth F, Israel A. Inhibition of the DNA-binding activity of Drosophila suppressor of hairless and of its human homolog, KBF2/RBP-Jκ, by direct protein– protein interaction with Drosophila hairless. Genes & Dev. 1994;8:2491–2503. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources