

Targeting histone deacetylase complexes via KRAB-zinc finger proteins: the PHD and bromodomains of KAP-1 form a cooperative unit that recruits a novel isoform of the Mi-2alpha subunit of NuRD
- PMID: 11230151
- PMCID: PMC312636
- DOI: 10.1101/gad.869501
Targeting histone deacetylase complexes via KRAB-zinc finger proteins: the PHD and bromodomains of KAP-1 form a cooperative unit that recruits a novel isoform of the Mi-2alpha subunit of NuRD
Abstract
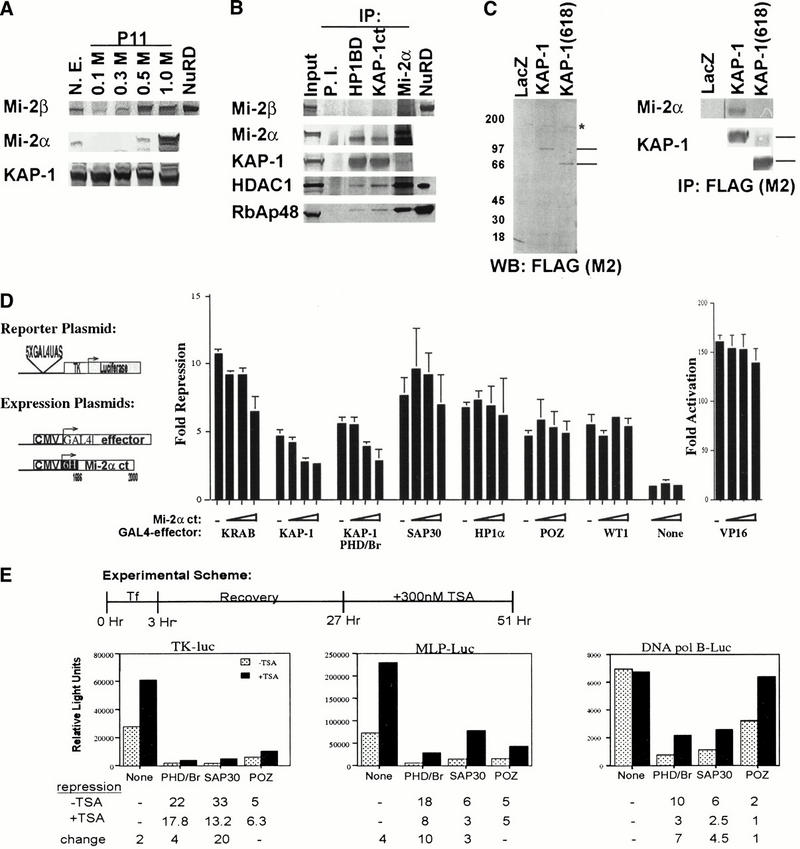
Macromolecular complexes containing histone deacetylase and ATPase activities regulate chromatin dynamics and are vitally responsible for transcriptional gene silencing in eukaryotes. The mechanisms that target these assemblies to specific loci are not as well understood. We show that the corepressor KAP-1, via its PHD (plant homeodomain) and bromodomain, links the superfamily of Krüppel associated box (KRAB) zinc finger proteins (ZFP) to the NuRD complex. We demonstrate that the tandem PHD finger and bromodomain of KAP-1, an arrangement often found in cofactor proteins but functionally ill-defined, form a cooperative unit that is required for transcriptional repression. Substitution of highly related PHD fingers or bromodomains failed to restore repression activity, suggesting high specificity in their cooperative function. Moreover, single amino acid substitutions in either the bromodomain or PHD finger, including ones that mimic disease-causing mutations in the hATRX PHD finger, abolish repression. A search for effectors of this repression function yielded a novel isoform of the Mi-2alpha protein, an integral component of the NuRD complex. Endogenous KAP-1 is associated with Mi-2alpha and other components of NuRD, and KAP-1-mediated silencing requires association with NuRD and HDAC activity. These data suggest the KRAB-ZFP superfamily of repressors functions to target the histone deacetylase and chromatin remodeling activities of the NuRD complex to specific gene promoters in vivo.
Figures



Similar articles
-
Reconstitution of the KRAB-KAP-1 repressor complex: a model system for defining the molecular anatomy of RING-B box-coiled-coil domain-mediated protein-protein interactions.J Mol Biol. 2000 Feb 4;295(5):1139-62. doi: 10.1006/jmbi.1999.3402. J Mol Biol. 2000. PMID: 10653693
-
A common DNA-binding site for SZF1 and the BRCA1-associated zinc finger protein, ZBRK1.Cancer Res. 2002 Jul 1;62(13):3773-81. Cancer Res. 2002. PMID: 12097288
-
Developmental roles of the Mi-2/NURD-associated protein p66 in Drosophila.Genetics. 2005 Apr;169(4):2087-100. doi: 10.1534/genetics.104.034595. Epub 2005 Feb 3. Genetics. 2005. PMID: 15695365 Free PMC article.
-
NuRD and SIN3 histone deacetylase complexes in development.Trends Genet. 2000 Aug;16(8):351-6. doi: 10.1016/s0168-9525(00)02066-7. Trends Genet. 2000. PMID: 10904264 Review.
-
Mi-2/NuRD: multiple complexes for many purposes.Biochim Biophys Acta. 2004 Mar 15;1677(1-3):52-7. doi: 10.1016/j.bbaexp.2003.10.010. Biochim Biophys Acta. 2004. PMID: 15020045 Review.
Cited by
-
ZNF91 deletion in human embryonic stem cells leads to ectopic activation of SVA retrotransposons and up-regulation of KRAB zinc finger gene clusters.Genome Res. 2021 Apr;31(4):551-563. doi: 10.1101/gr.265348.120. Epub 2021 Mar 15. Genome Res. 2021. PMID: 33722937 Free PMC article.
-
MBD3 expression and DNA binding patterns are altered in a rat model of temporal lobe epilepsy.Sci Rep. 2016 Sep 21;6:33736. doi: 10.1038/srep33736. Sci Rep. 2016. PMID: 27650712 Free PMC article.
-
Genetic Studies on Mammalian DNA Methyltransferases.Adv Exp Med Biol. 2022;1389:111-136. doi: 10.1007/978-3-031-11454-0_5. Adv Exp Med Biol. 2022. PMID: 36350508 Free PMC article.
-
KRAB domain of ZFP568 disrupts TRIM28-mediated abnormal interactions in cancer cells.NAR Cancer. 2020 Jun;2(2):zcaa007. doi: 10.1093/narcan/zcaa007. Epub 2020 May 18. NAR Cancer. 2020. PMID: 32743551 Free PMC article.
-
TRIM28 functions as the SUMO E3 ligase for PCNA in prevention of transcription induced DNA breaks.Proc Natl Acad Sci U S A. 2020 Sep 22;117(38):23588-23596. doi: 10.1073/pnas.2004122117. Epub 2020 Sep 8. Proc Natl Acad Sci U S A. 2020. PMID: 32900933 Free PMC article.
References
-
- Aasland R, Gibson TJ, Stewart AF. The PHD finger: Implications for chromatin-mediated transcriptional regulation. Trends Biochem Sci. 1995;20:56–59. - PubMed
-
- Agata Y, Matsuda E, Shimizu A. Two novel Kruppel-associated box-containing zinc-finger proteins, KRAZ1 and KRAZ2, repress transcription through functional interaction with the corepressor KAP-1 (TIF1beta/KRIP-1) J Biol Chem. 1999;274:16412–16422. - PubMed
-
- Bjorses P, Halonen M, Palvimo JJ, Kolmer M, Aaltonen J, Ellonen P, Perheentupa J, Ulmanen I, Peltonen L. Mutations in the AIRE gene: Effects on subcellular location and transactivation function of the autoimmune polyendocrinopathy–candidiasis–ectodermal dystrophy protein. Am J Hum Genet. 2000;66:378–392. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous