

Identification of human epidermal differentiation complex (EDC)-encoded genes by subtractive hybridization of entire YACs to a gridded keratinocyte cDNA library
- PMID: 11230159
- PMCID: PMC311024
- DOI: 10.1101/gr.114801
Identification of human epidermal differentiation complex (EDC)-encoded genes by subtractive hybridization of entire YACs to a gridded keratinocyte cDNA library
Abstract
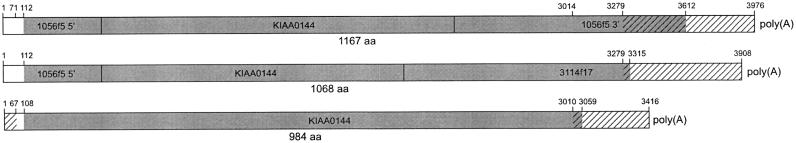
The epidermal differentiation complex (EDC) comprises a large number of genes that are of crucial importance for the maturation of the human epidermis. So far, 27 genes of 3 related families encoding structural as well as regulatory proteins have been mapped within a 2-Mb region on chromosome 1q21. Here we report on the identification of 10 additional EDC genes by a powerful subtractive hybridization method using entire YACs (950_e_2 and 986_e_10) to screen a gridded human keratinocyte cDNA library. Localization of the detected cDNA clones has been established on a long-range restriction map covering more than 5 Mb of this genomic region. The genes encode cytoskeletal tropomyosin TM30nm (TPM3), HS1-binding protein Hax-1 (HAX1), RNA-specific adenosine deaminase (ADAR1), the 34/67-kD laminin receptor (LAMRL6), and the 26S proteasome subunit p31 (PSMD8L), as well as five hitherto uncharacterized proteins (NICE-1, NICE-2, NICE-3, NICE-4, and NICE-5). The nucleotide sequences and putative ORFs of the EDC genes identified here revealed no homology with any of the established EDC gene families. Whereas database searches revealed that NICE-3, NICE-4, and NICE-5 were expressed in many tissues, no EST or gene-specific sequence was found for NICE-2. Expression of NICE-1 was up-regulated in differentiated keratinocytes, pointing to its relevance for the terminal differentiation of the epidermis. The newly identified EDC genes are likely to provide further insights into epidermal differentiation and they are potential candidates to be involved in skin diseases and carcinogenesis that are associated with this region of chromosome 1. Moreover, the extended integrated map of the EDC, including the polymorphic sequence tag site (STS) markers D1S1664, D1S2346, and D1S305, will serve as a valuable tool for linkage analyses.
Figures







Similar articles
-
Genetic analysis of the epidermal differentiation complex (EDC) on human chromosome 1q21: chromosomal orientation, new markers, and a 6-Mb YAC contig.Genomics. 1996 Nov 1;37(3):295-302. doi: 10.1006/geno.1996.0563. Genomics. 1996. PMID: 8938441
-
Isolation of cDNAs from the Cri-du-chat critical region by direct screening of a chromosome 5-specific cDNA library.Genome Res. 1997 Feb;7(2):118-27. doi: 10.1101/gr.7.2.118. Genome Res. 1997. PMID: 9049630
-
Comparison of DNA Methylation and Expression Pattern of S100 and Other Epidermal Differentiation Complex Genes in Differentiating Keratinocytes.J Cell Biochem. 2016 May;117(5):1092-8. doi: 10.1002/jcb.25392. Epub 2015 Oct 14. J Cell Biochem. 2016. PMID: 26443750
-
The human epidermal differentiation complex: cornified envelope precursors, S100 proteins and the 'fused genes' family.Exp Dermatol. 2012 Sep;21(9):643-9. doi: 10.1111/j.1600-0625.2012.01472.x. Epub 2012 Apr 16. Exp Dermatol. 2012. PMID: 22507538 Review.
-
Update on the epidermal differentiation complex.Front Biosci (Landmark Ed). 2012 Jan 1;17(4):1517-32. doi: 10.2741/4001. Front Biosci (Landmark Ed). 2012. PMID: 22201818 Review.
Cited by
-
Peptidoglycan recognition proteins Pglyrp3 and Pglyrp4 are encoded from the epidermal differentiation complex and are candidate genes for the Psors4 locus on chromosome 1q21.Hum Genet. 2006 Mar;119(1-2):113-25. doi: 10.1007/s00439-005-0115-8. Epub 2005 Dec 17. Hum Genet. 2006. PMID: 16362825
-
Small proline-rich repeat 3 is a novel coordinator of PDGFRβ and integrin β1 crosstalk to augment proliferation and matrix synthesis by cardiac fibroblasts.FASEB J. 2020 Jun;34(6):7885-7904. doi: 10.1096/fj.201902815R. Epub 2020 Apr 16. FASEB J. 2020. PMID: 32297675 Free PMC article.
-
Suprabasin, a novel epidermal differentiation marker and potential cornified envelope precursor.J Biol Chem. 2002 Nov 22;277(47):45195-202. doi: 10.1074/jbc.M205380200. Epub 2002 Sep 12. J Biol Chem. 2002. PMID: 12228223 Free PMC article.
-
Certainty of S100 from Physiology to Pathology.J Clin Diagn Res. 2016 Jun;10(6):ZE10-5. doi: 10.7860/JCDR/2016/17949.8022. Epub 2016 Jun 1. J Clin Diagn Res. 2016. PMID: 27504432 Free PMC article. Review.
-
Autoantigen Discovery in the Hair Loss Disorder, Alopecia Areata: Implication of Post-Translational Modifications.Front Immunol. 2022 Jun 3;13:890027. doi: 10.3389/fimmu.2022.890027. eCollection 2022. Front Immunol. 2022. PMID: 35720384 Free PMC article. Review.
References
-
- Albertazzi E, Cajone F, Leone BE, Naguib RN, Lakshmi MS, Sherbet GV. Expression of metastasis-associated genes h-mts1 (S100A4) and nm23 in carcinoma of breast is related to disease progression. DNA Cell Biol. 1998;17:335–342. - PubMed
-
- Backendorf C, Hohl D. A common origin for cornified envelope proteins? Nat Genet. 1992;2:91. - PubMed
-
- Bignon C, Roux-Dosseto M, Zeigler ME, Mattei MG, Lissitzky JC, Wicha MS, Martin PM. Genomic analysis of the 67-kDa laminin receptor in normal and pathological tissues: Circumstantial evidence for retroposon features. Genomics. 1991;10:481–485. - PubMed
-
- Boultwood J, Fidler C, Soularue P, Strickson AJ, Kostrzewa M, Jaju RJ, Cotter FE, Fairweather N, Monaco AP, Muller U, et al. Novel genes mapping to the critical region of the 5q-syndrome. Genomics. 1997;45:88–96. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous