

Expression of hypoxia-inducible cell-surface transmembrane carbonic anhydrases in human cancer
- PMID: 11238039
- PMCID: PMC1850340
- DOI: 10.1016/S0002-9440(10)64038-2
Expression of hypoxia-inducible cell-surface transmembrane carbonic anhydrases in human cancer
Abstract
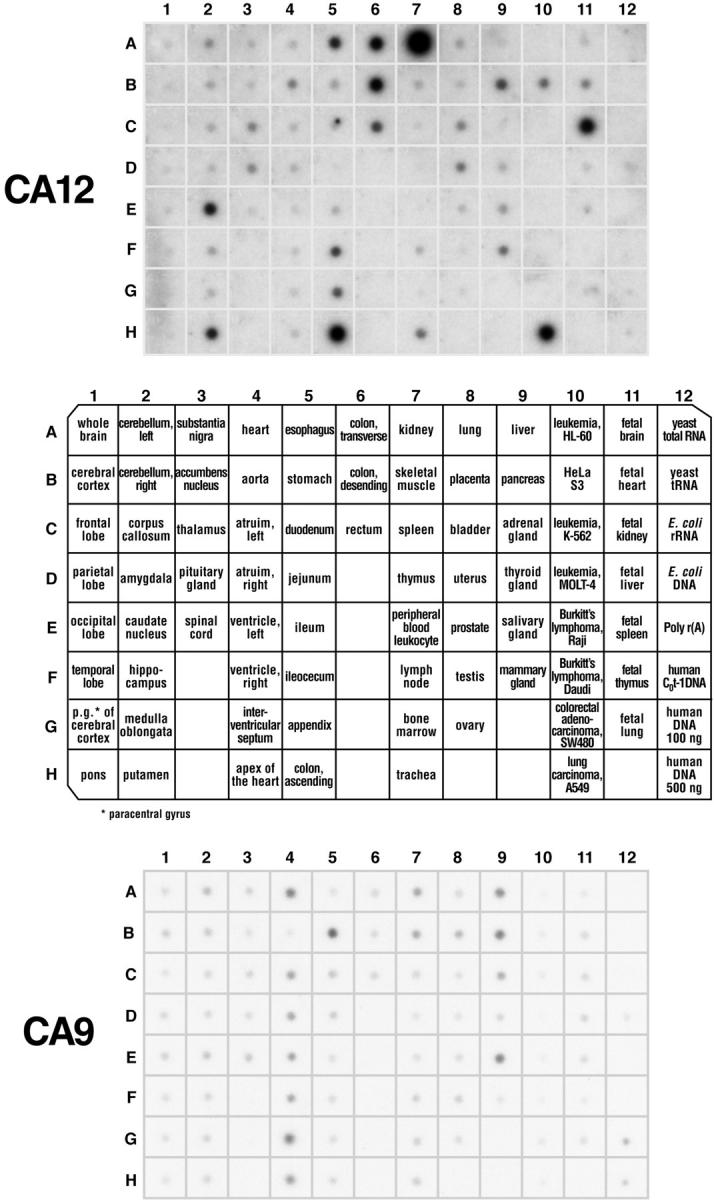
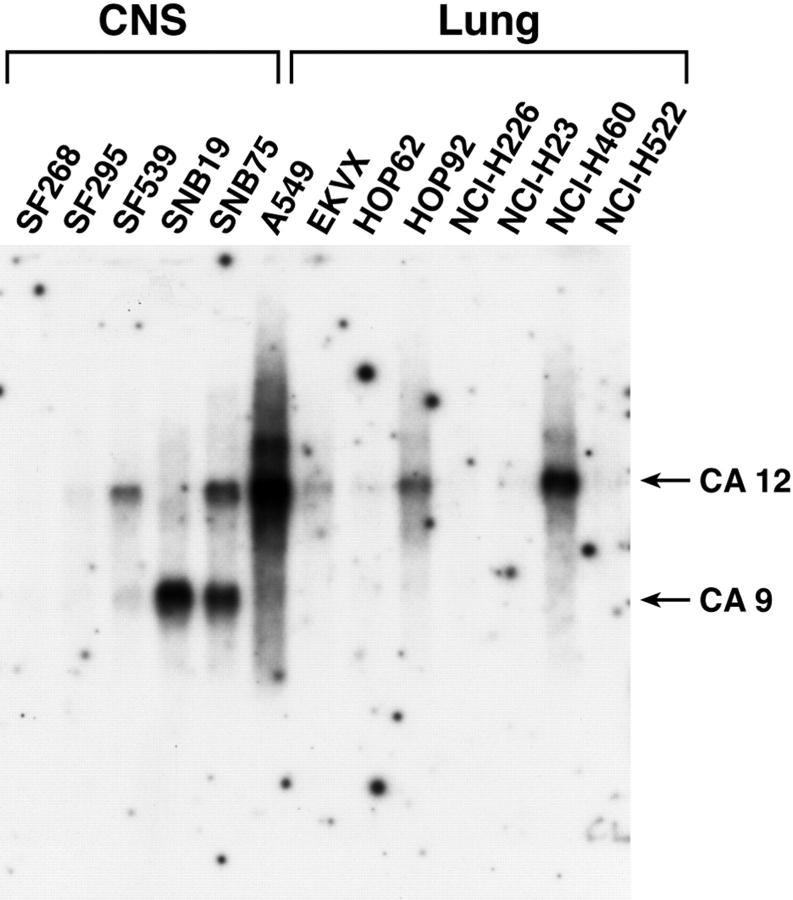
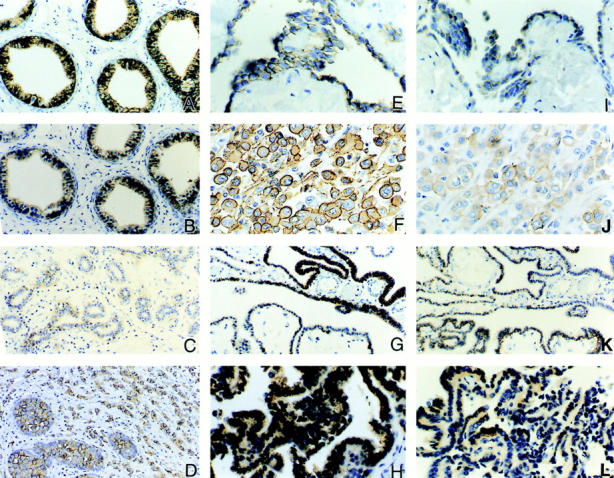
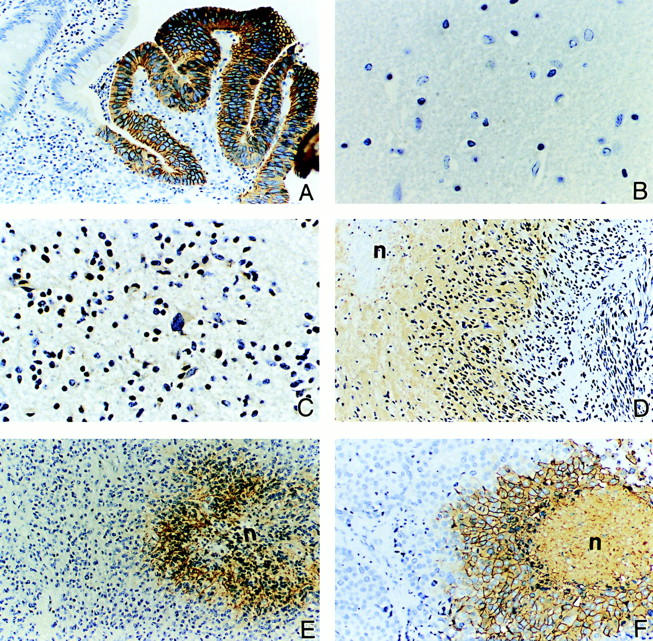
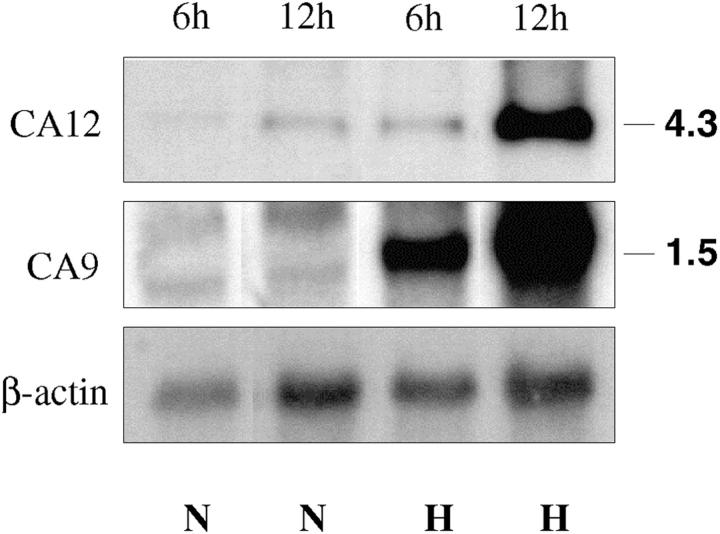
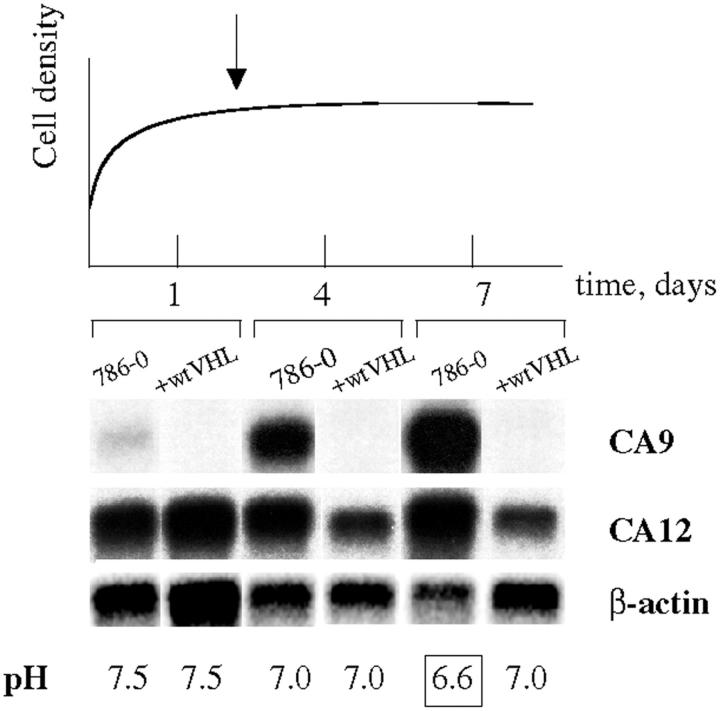
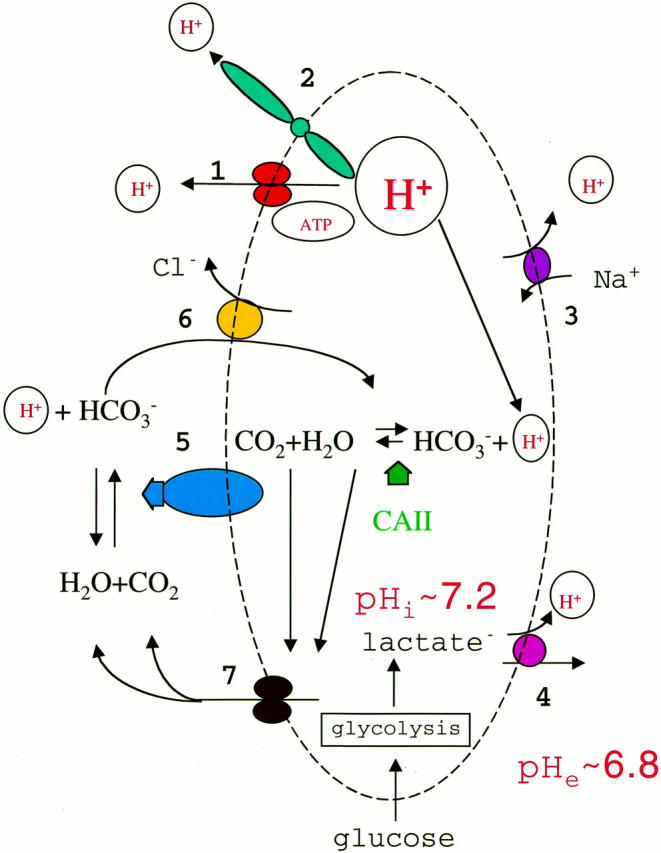
An acidic extracellular pH is a fundamental property of the malignant phenotype. In von Hippel-Lindau (VHL)-defective tumors the cell surface transmembrane carbonic anhydrase (CA) CA9 and CA12 genes are overexpressed because of the absence of pVHL. We hypothesized that these enzymes might be involved in maintaining the extracellular acidic pH in tumors, thereby providing a conducive environment for tumor growth and spread. Using Northern blot analysis and immunostaining with specific antibodies we analyzed the expression of CA9 and CA12 genes and their products in a large sample of cancer cell lines, fresh and archival tumor specimens, and normal human tissues. Expression was also analyzed in cultured cells under hypoxic conditions. Expression of CA IX and CA XII in normal adult tissues was detected only in highly specialized cells and for most tissues their expression did not overlap. Analysis of RNA samples isolated from 87 cancer cell lines and 18 tumors revealed high-to-moderate levels of expression of CA9 and CA12 in multiple cancers. Immunohistochemistry revealed high-to-moderate expression of these enzymes in various normal tissues and multiple common epithelial tumor types. The immunostaining was seen predominantly on the cell surface membrane. The expression of both genes was markedly induced under hypoxic conditions in tumors and cultured tumor cells. We conclude that the cell surface trans-membrane carbonic anhydrases CA IX and CA XII are overexpressed in many tumors suggesting that this is a common feature of cancer cells that may be required for tumor progression. These enzymes may contribute to the tumor microenvironment by maintaining extracellular acidic pH and helping cancer cells grow and metastasize. Our studies show an important causal link between hypoxia, extracellular acidification, and induction or enhanced expression of these enzymes in human tumors.
Figures

References
-
- Semenza GL: Hypoxia, clonal selection, and the role of HIF-1 in tumor progression. Crit Rev Biochem Mol Biol 2000, 35:71-103 - PubMed
-
- Hanahan D, Folkman J: Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell 1996, 86:353-364 - PubMed
-
- Warburg O: The Metabolism of Tumours. 1930. Constable, London
-
- Dang CV, Semenza GL: Oncogenic alterations of metabolism. Trends Biochem Sci 1999, 24:68-72 - PubMed
-
- Stubbs M, McSheehy PM, Griffiths JR, Bashford CL: Causes and consequences of tumour acidity and implications for treatment. Mol Med Today 2000, 6:15-19 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
