

The localization of human cyclins B1 and B2 determines CDK1 substrate specificity and neither enzyme requires MEK to disassemble the Golgi apparatus
- PMID: 11238451
- PMCID: PMC2198800
- DOI: 10.1083/jcb.152.5.945
The localization of human cyclins B1 and B2 determines CDK1 substrate specificity and neither enzyme requires MEK to disassemble the Golgi apparatus
Abstract
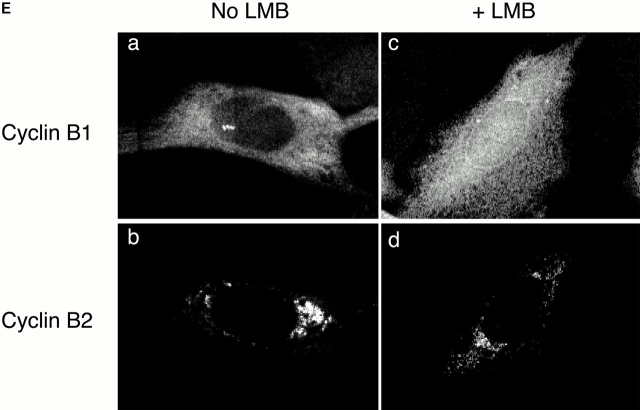
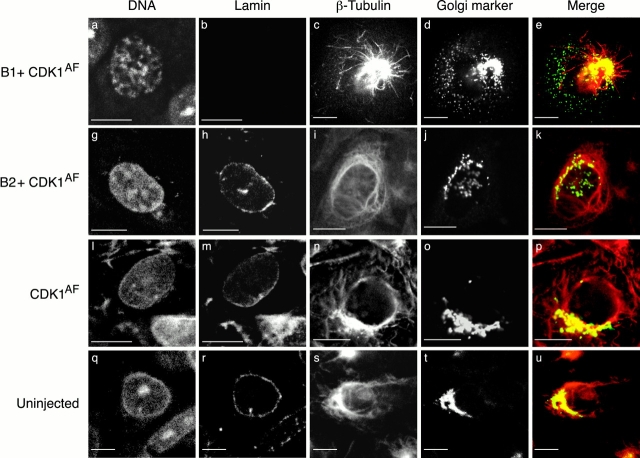
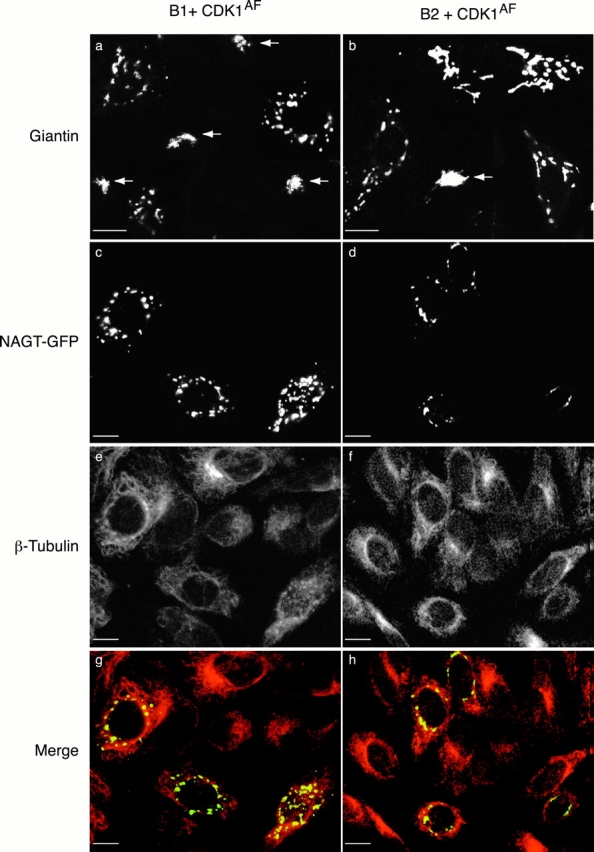
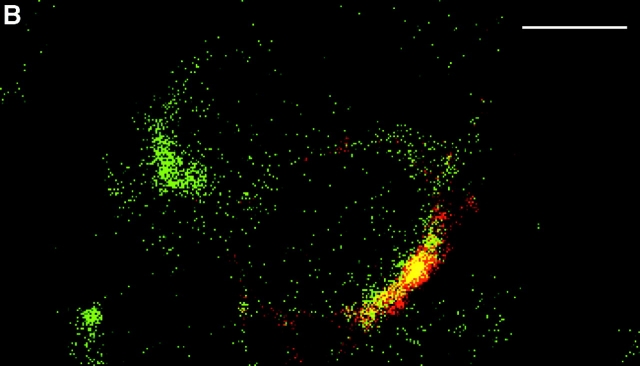
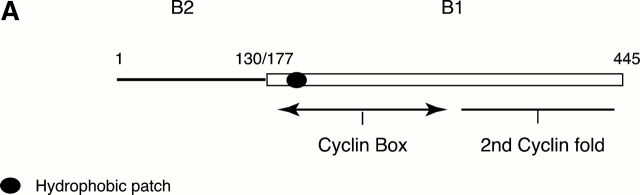
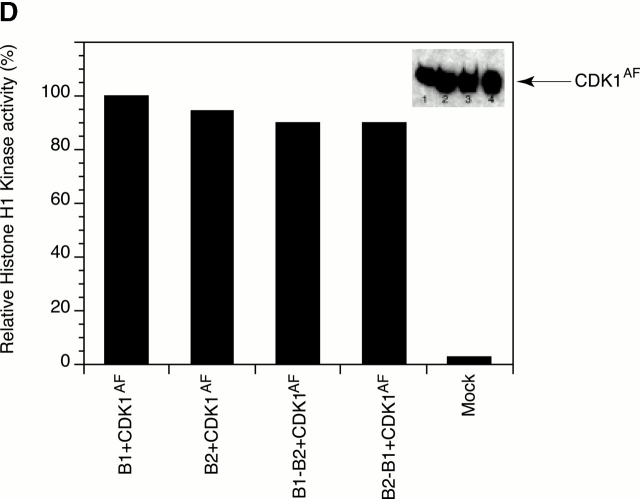
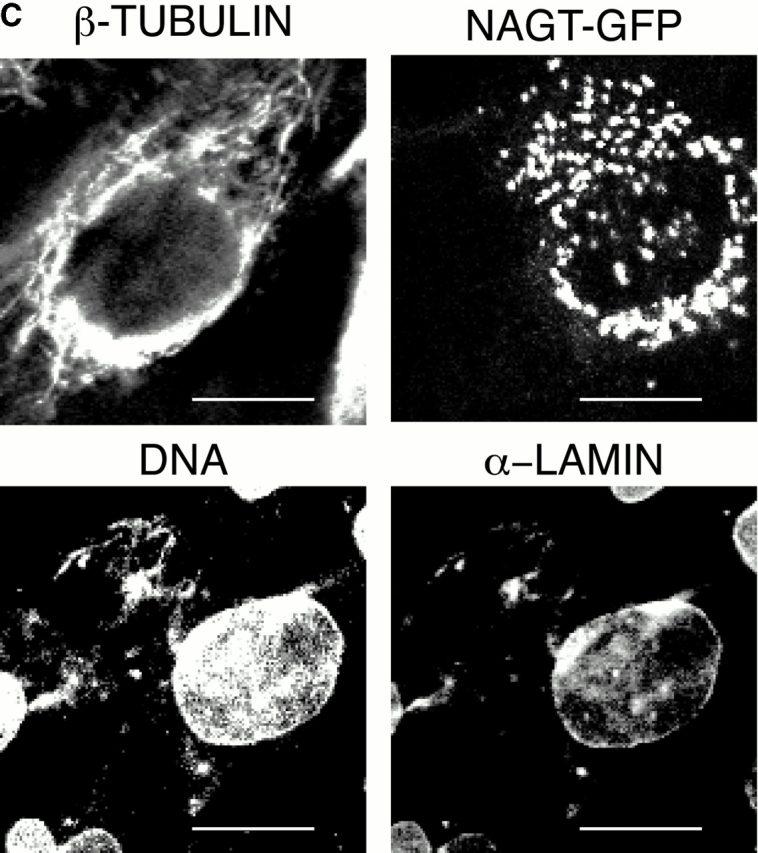
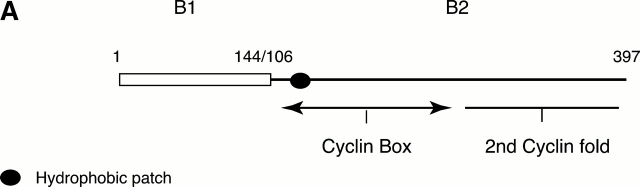
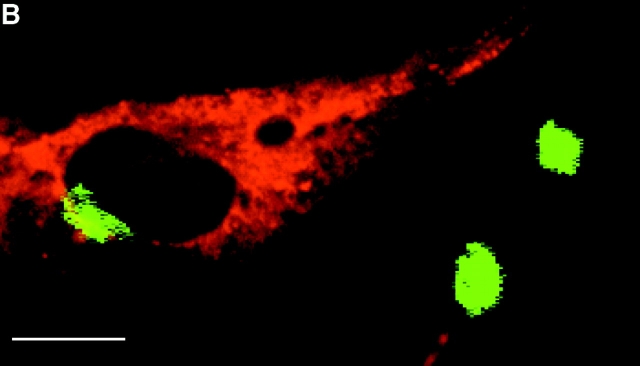
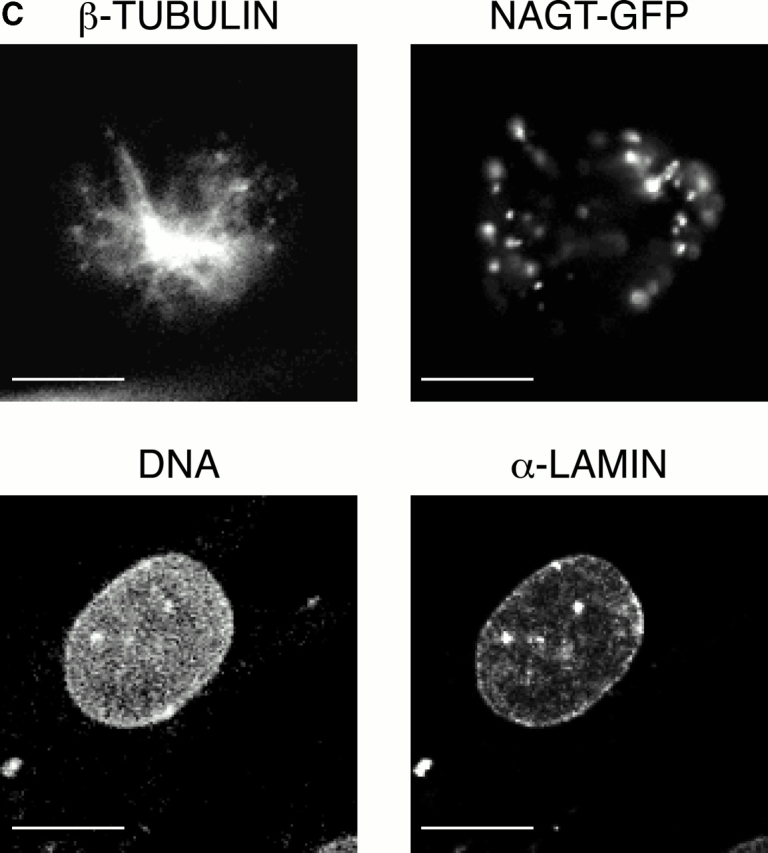
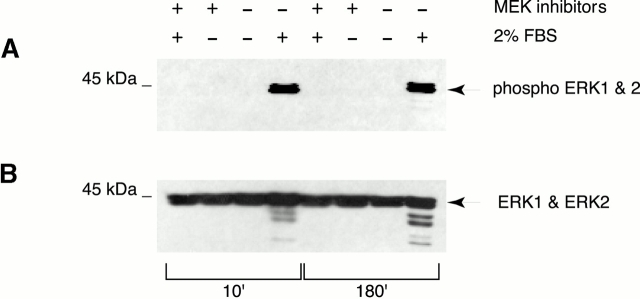
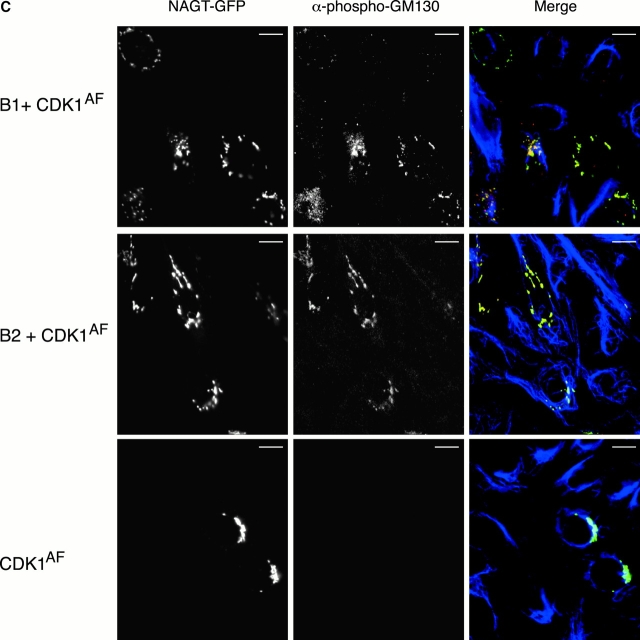
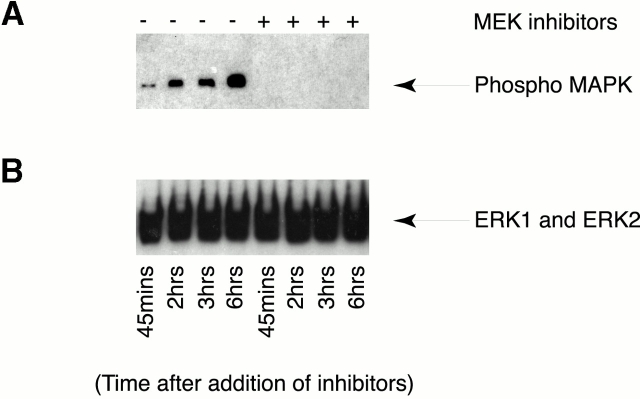
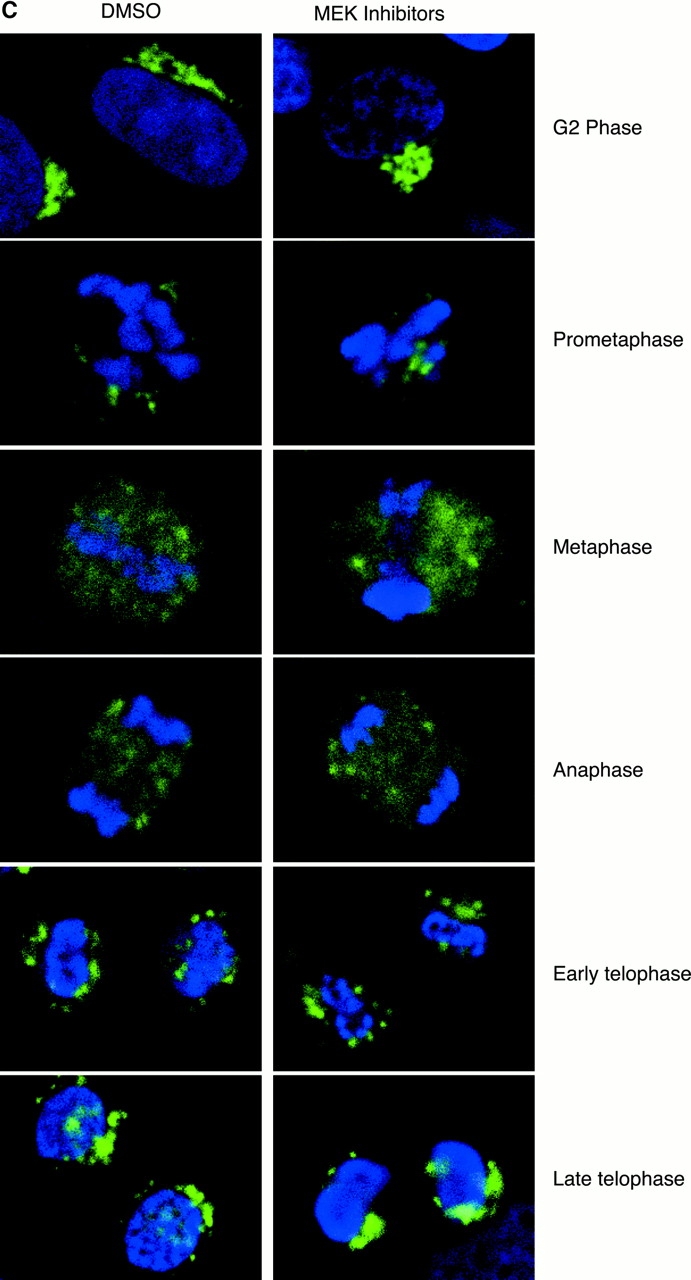
In this paper, we show that substrate specificity is primarily conferred on human mitotic cyclin-dependent kinases (CDKs) by their subcellular localization. The difference in localization of the B-type cyclin-CDKs underlies the ability of cyclin B1-CDK1 to cause chromosome condensation, reorganization of the microtubules, and disassembly of the nuclear lamina and of the Golgi apparatus, while it restricts cyclin B2-CDK1 to disassembly of the Golgi apparatus. We identify the region of cyclin B2 responsible for its localization and show that this will direct cyclin B1 to the Golgi apparatus and confer upon it the more limited properties of cyclin B2. Equally, directing cyclin B2 to the cytoplasm with the NH(2) terminus of cyclin B1 confers the broader properties of cyclin B1. Furthermore, we show that the disassembly of the Golgi apparatus initiated by either mitotic cyclin-CDK complex does not require mitogen-activated protein kinase kinase (MEK) activity.
Figures
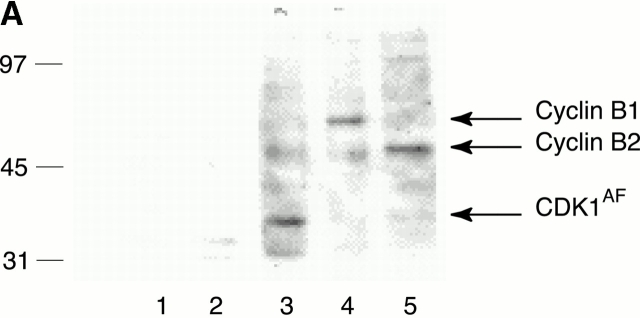
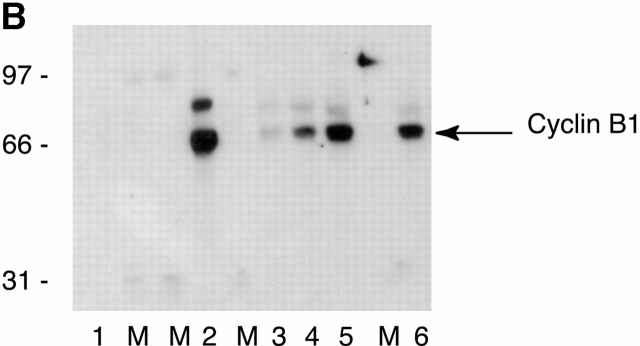
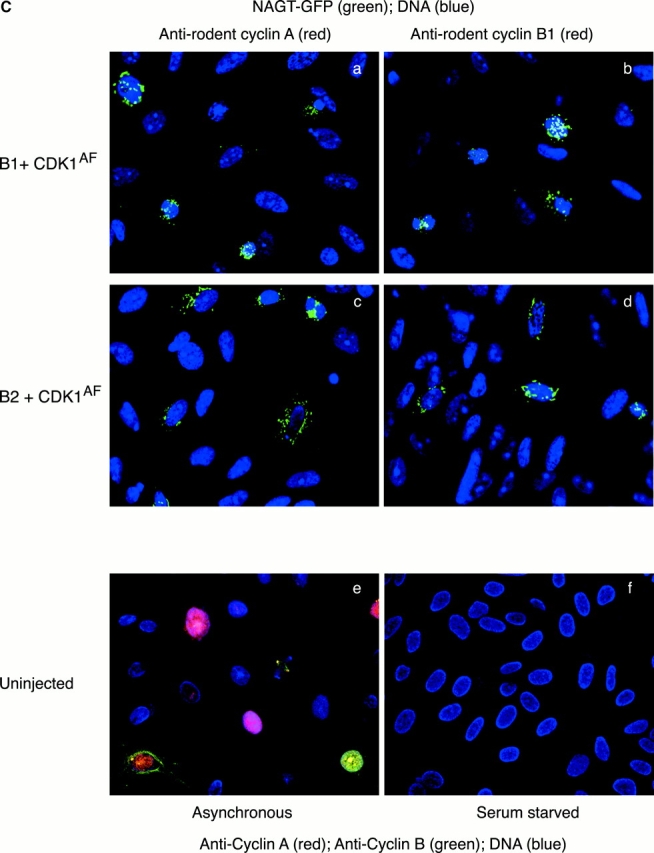
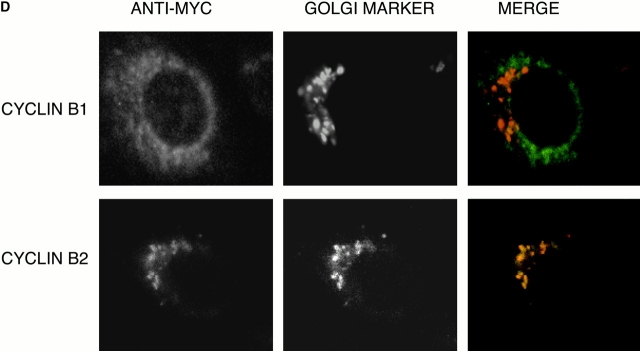
References
-
- Acharya U., Mallabiabarrena A., Acharya J.K., Malhotra V. Signaling via mitogen-activated protein kinase kinase (MEK1) is required for Golgi fragmentation during mitosis. Cell. 1998;92:183–192. - PubMed
-
- Chapman D.L., Wolgemuth D.J. Isolation of the murine cyclin B2 cDNA and characterization of the lineage and temporal specificity of expression of the B1 and B2 cyclins during oogenesis, spermatogenesis and early embryogenesis. Development. 1993;118:229–240. - PubMed
-
- Chen J., Jackson P.K., Kirschner M.W., Dutta A. Separate domains of p21 involved in the inhibition of cdk kinase and PCNA. Nature. 1995;374:386–388. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
