

CD28-independent induction of experimental autoimmune encephalomyelitis
- PMID: 11238558
- PMCID: PMC199425
- DOI: 10.1172/JCI11220
CD28-independent induction of experimental autoimmune encephalomyelitis
Abstract
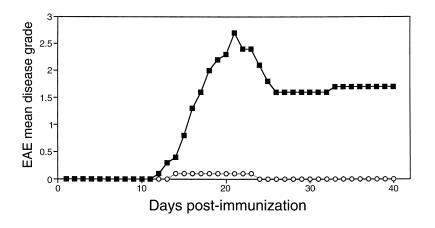
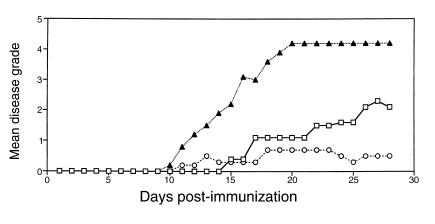
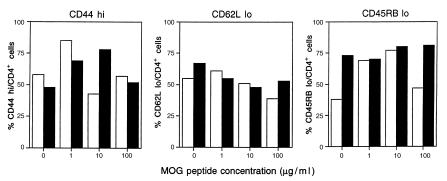
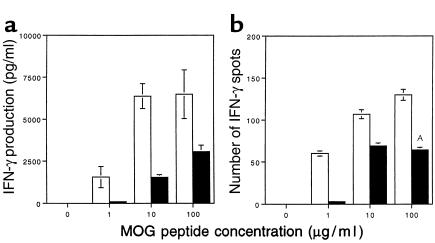
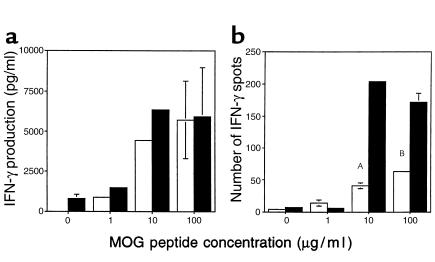
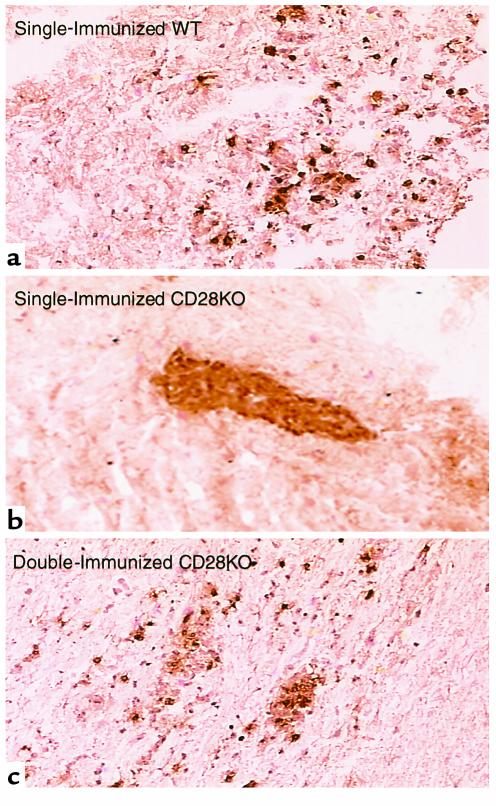
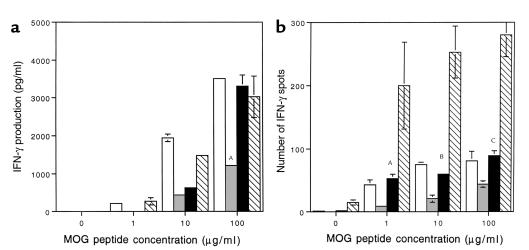
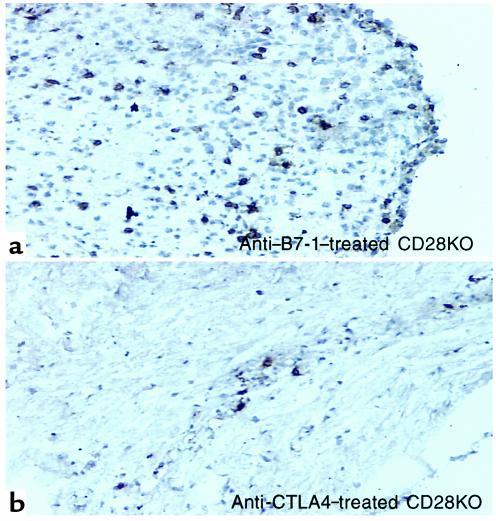
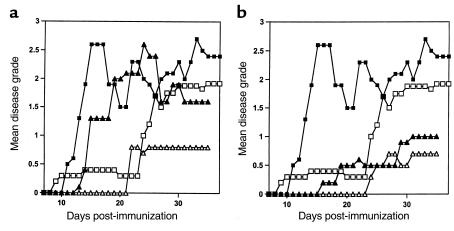
Experimental autoimmune encephalomyelitis (EAE) is a T cell-mediated disease initiated by antigen-specific CD4(+) T cells. Signaling through CD28 is a critical second signal for activation of T cells, and CD28 knockout (CD28KO) mice have been reported to be resistant to induction of EAE. We now report that CD28KO mice have no intrinsic defect in mediating disease, because they developed EAE after passive transfer of primed T cells. After immunization, peripheral T cells from CD28KO mice were primed and developed memory phenotype, but had decreased antigen-specific IFN-gamma production as compared with cells from wild-type (WT) animals. Reimmunization of CD28KO mice brought out clinical disease and increased IFN-gamma production in vitro. Pathologically, there were cellular infiltrates in the central nervous system, in contrast to single-immunized mice. We show furthermore that blocking B7-1 or CTLA4, but not B7-2, in CD28KO mice induces disease after a single immunization. Thus, EAE can be induced in animals lacking CD28-dependent costimulation, suggesting that alternative costimulatory pathways were used. Blocking the OX40-OX40L costimulatory pathway differentially affected disease induction in CD28KO mice as compared with WT controls. Our data show that EAE may develop in the absence of CD28 T-cell costimulation. These findings have implications for therapies aimed at blocking costimulatory signals in autoimmune diseases.
Figures
Similar articles
-
Critical involvement of OX40 ligand signals in the T cell priming events during experimental autoimmune encephalomyelitis.J Immunol. 2001 Sep 1;167(5):2991-9. doi: 10.4049/jimmunol.167.5.2991. J Immunol. 2001. PMID: 11509650
-
Studies in B7-deficient mice reveal a critical role for B7 costimulation in both induction and effector phases of experimental autoimmune encephalomyelitis.J Exp Med. 1999 Sep 6;190(5):733-40. doi: 10.1084/jem.190.5.733. J Exp Med. 1999. PMID: 10477557 Free PMC article.
-
Differential requirements of naïve and memory T cells for CD28 costimulation in autoimmune pathogenesis.Histol Histopathol. 1999 Oct;14(4):1269-76. doi: 10.14670/HH-14.1269. Histol Histopathol. 1999. PMID: 10506942 Review.
-
B7 interactions with CD28 and CTLA-4 control tolerance or induction of mucosal inflammation in chronic experimental colitis.J Immunol. 2001 Aug 1;167(3):1830-8. doi: 10.4049/jimmunol.167.3.1830. J Immunol. 2001. PMID: 11466409
-
B7-mediated costimulation can either provoke or prevent clinical manifestations of experimental allergic encephalomyelitis.Immunol Res. 1995;14(3):189-99. doi: 10.1007/BF02918216. Immunol Res. 1995. PMID: 8778209 Review.
Cited by
-
Critical role of the programmed death-1 (PD-1) pathway in regulation of experimental autoimmune encephalomyelitis.J Exp Med. 2003 Jul 7;198(1):71-8. doi: 10.1084/jem.20022119. J Exp Med. 2003. PMID: 12847138 Free PMC article.
-
Targeting PGLYRP1 in cancer and autoimmunity.Nat Immunol. 2023 Nov;24(11):1785-1786. doi: 10.1038/s41590-023-01649-0. Nat Immunol. 2023. PMID: 37828380 No abstract available.
-
Regulation of arginase I activity and expression by both PD-1 and CTLA-4 on the myeloid-derived suppressor cells.Cancer Immunol Immunother. 2009 May;58(5):687-97. doi: 10.1007/s00262-008-0591-5. Epub 2008 Oct 1. Cancer Immunol Immunother. 2009. PMID: 18828017 Free PMC article.
-
CD40-mediated signalling influences trafficking, T-cell receptor expression, and T-cell pathogenesis, in the NOD model of type 1 diabetes.Immunology. 2017 Oct;152(2):243-254. doi: 10.1111/imm.12761. Epub 2017 Jun 19. Immunology. 2017. PMID: 28542921 Free PMC article.
-
Effect of targeted disruption of STAT4 and STAT6 on the induction of experimental autoimmune encephalomyelitis.J Clin Invest. 2001 Sep;108(5):739-47. doi: 10.1172/JCI12563. J Clin Invest. 2001. PMID: 11544280 Free PMC article.
References
-
- Mueller DL, Jenkins MK, Schwartz RH. Clonal expansion versus functional clonal inactivation: a costimulatory signalling pathway determines the outcome of T cell antigen receptor occupancy. Annu Rev Immunol. 1989;7:445–480. - PubMed
-
- Schwartz RH. A cell culture model for T lymphocyte clonal anergy. Science. 1990;248:1349–1356. - PubMed
-
- Walunas TL, et al. CTLA-4 can function as a negative regulator of T cell activation. Immunity. 1994;1:405–413. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
