

Epitope mapping porcine reproductive and respiratory syndrome virus by phage display: the nsp2 fragment of the replicase polyprotein contains a cluster of B-cell epitopes
- PMID: 11238854
- PMCID: PMC114121
- DOI: 10.1128/JVI.75.7.3277-3290.2001
Epitope mapping porcine reproductive and respiratory syndrome virus by phage display: the nsp2 fragment of the replicase polyprotein contains a cluster of B-cell epitopes
Abstract
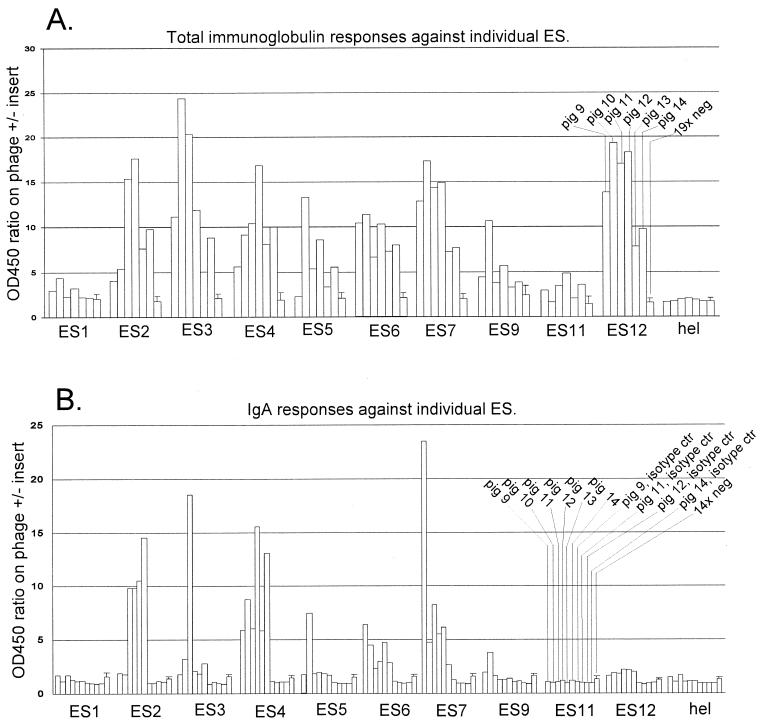
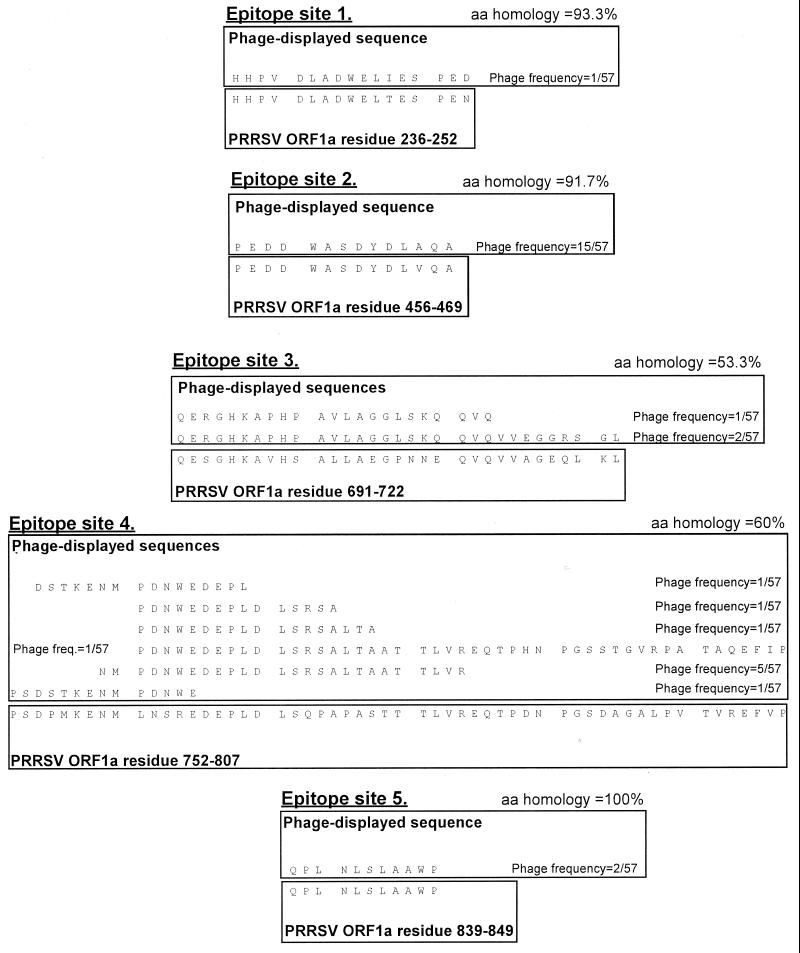
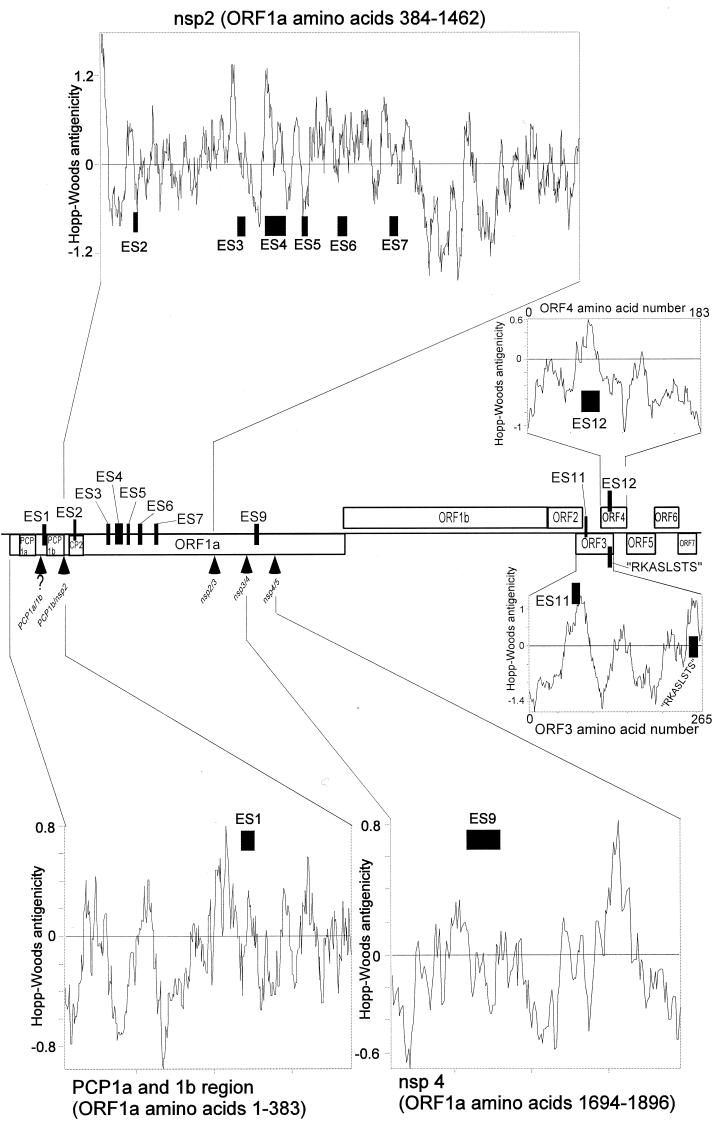
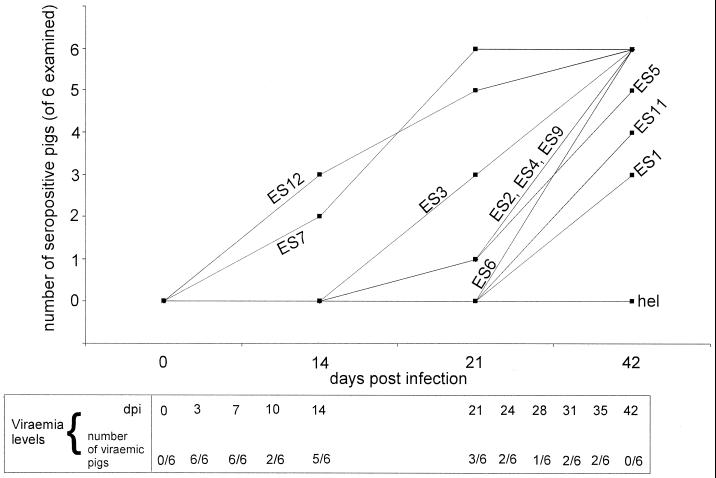
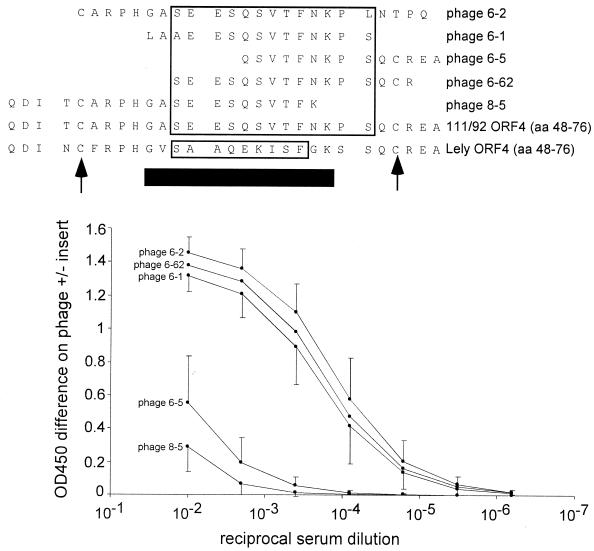
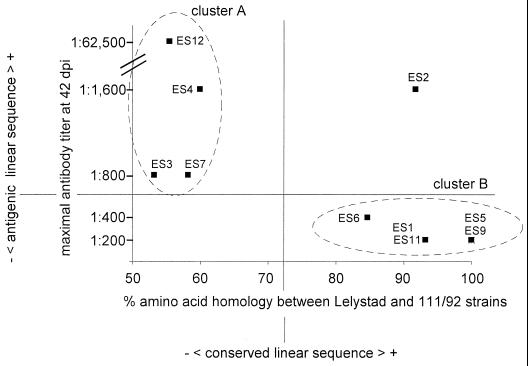
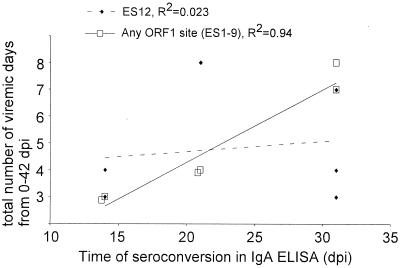
We screened phage display libraries of porcine reproductive and respiratory syndrome virus (PRRSV) protein fragments with sera from experimentally infected pigs to identify linear B-cell epitopes that are commonly recognized during infection in vivo. We identified 10 linear epitope sites (ES) 11 to 53 amino acids in length. In the replicase polyprotein, a total of eight ES were identified, six of which localized to the Nsp2 replicase polyprotein processing end product. In the structural proteins, a total of two ES were identified, in the ORF3 and ORF4 minor envelope glycoproteins. The ORF4 ES was previously identified by monoclonal antibody mapping (J. J. M. Meulenberg, A. P. van Nieuwstadt, A. van Essen-Zandenbergen, and J. P. M. Langeveld, J. Virol. 71:6061-6067, 1997), but its immunogenicity had not been examined in pigs. We found that six experimentally PRRSV-infected pigs consistently had very high antibody titers against the ORF4 ES. In some animals, sera diluted 1:62,500 still gave weak positive enzyme immunoassay reactivity against the ORF4 ES. This hitherto unrecognized immunodominance likely caused phages displaying the ORF4 ES to outcompete phages displaying other ES during library screening with porcine sera and accounted for our failure to identify more than two ES in the structural genes of PRRSV. Genetic analysis showed that variable ES were also the most immunogenic in vivo. Serological analysis indicated differences in the immunoglobulin A responses between short-term and longer-term viremic pigs towards some ES. The implications of these findings for PRRSV diagnostics and immunopathogenesis are discussed.
Figures

References
-
- Bøtner A, Nielsen J, Bille-Hansen V. Isolation of porcine reproductive and respiratory syndrome (PRRS) virus in a Danish swine herd and experimental infection of pregnant gilts with the virus. Vet Microbiol. 1994;40:351–360. - PubMed
-
- Bøtner A, Strandbygaard B, Sørensen K J, Have P, Madsen K G, Smedegaard Madsen E, Alexandersen S. Appearance of acute PRRS-like symptoms in sow herds after vaccination with a modified live PRRS vaccine. Vet Rec. 1997;141:497–499. - PubMed
-
- Christopher-Hennings J, Nelson E A, Hines R J, Nelson J K, Swenson S L, Zimmerman J J, Chase C L, Yaeger M J, Benfield D A. Persistence of porcine reproductive and respiratory syndrome virus in serum and semen of abult boars. J Vet Diagn Investig. 1995;7:456–464. - PubMed
-
- Cleveland S M, Buratti E, Jones T D, North P, Baralle F, McLain L, McInerney T, Durrani Z, Dimmock N J. Immunogenic and antigenic dominance of a nonneutralizing epitope over a highly conserved neutralizing epitope in the gp41 envelope glycoprotein of human immunodeficiency virus type 1: its deletion leads to a strong neutralizing response. Virology. 2000;266:66–78. - PubMed
-
- Day L A. Conformations of single-stranded DNA and coat protein in fd bacteriophage as revealed by ultraviolet absorption spectroscopy. J Mol Biol. 1969;39:265–277. - PubMed
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
