

Zygotic regulation of maternal cyclin A1 and B2 mRNAs
- PMID: 11238903
- PMCID: PMC86712
- DOI: 10.1128/MCB.21.5.1662-1671.2001
Zygotic regulation of maternal cyclin A1 and B2 mRNAs
Abstract
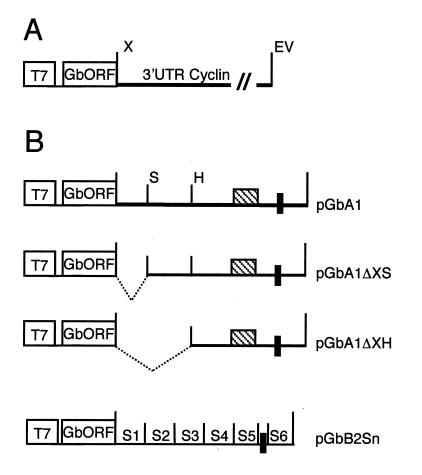
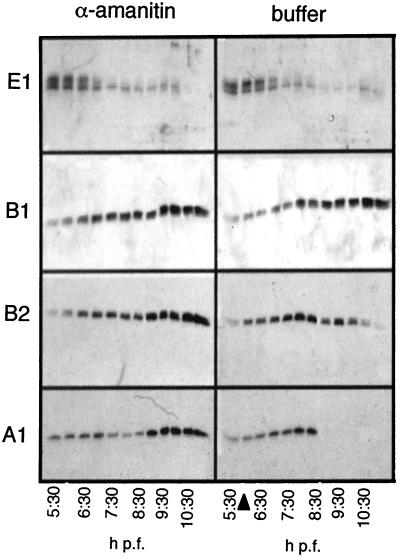
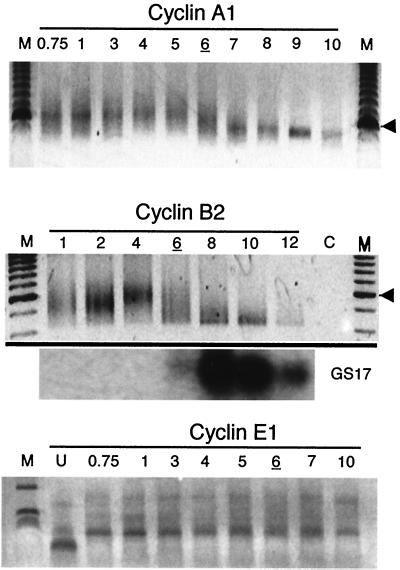
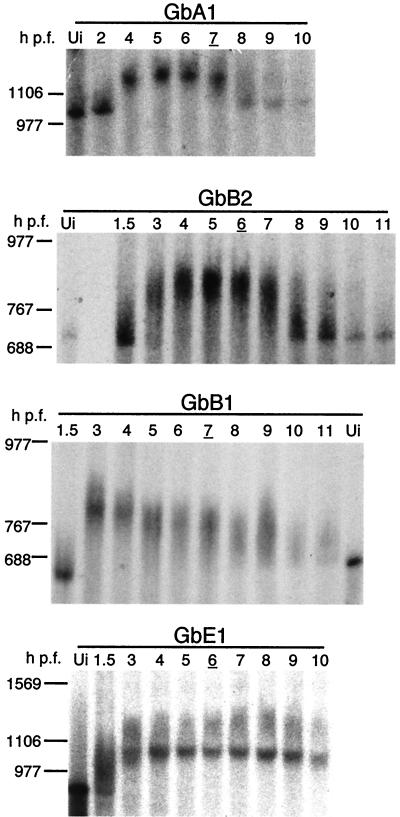
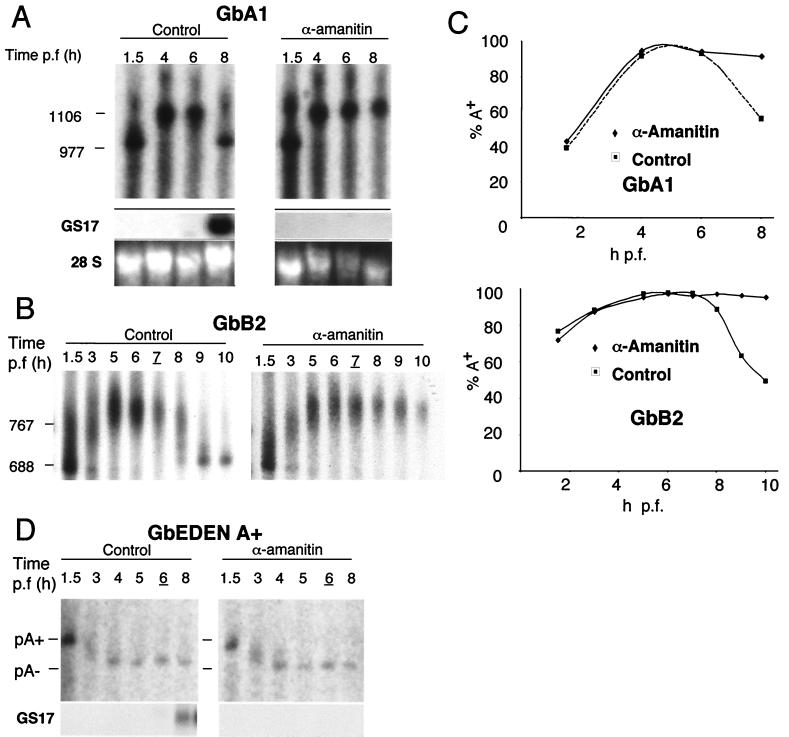
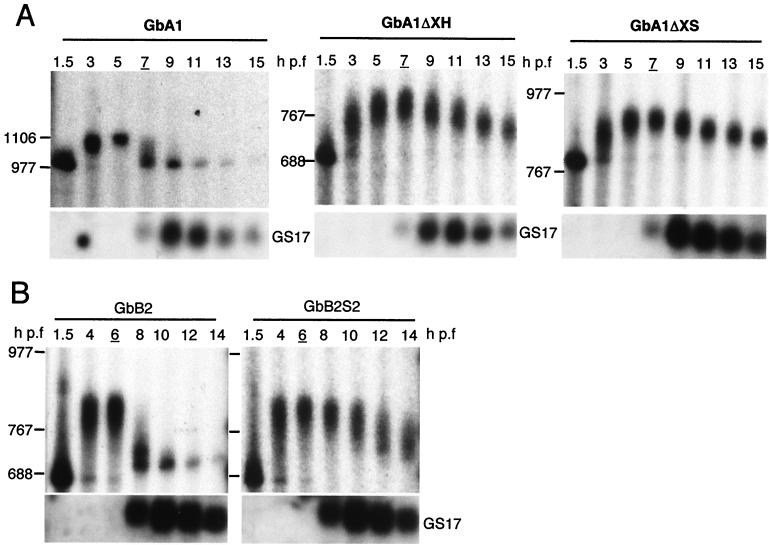
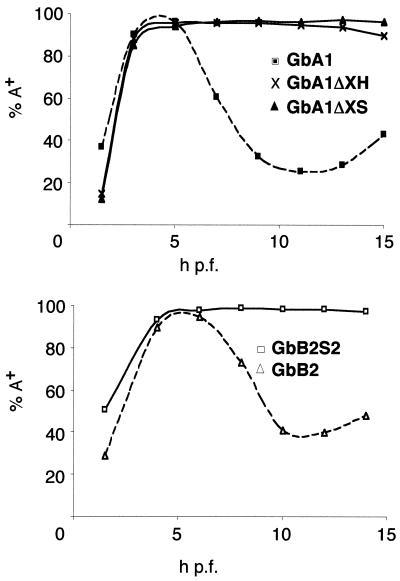
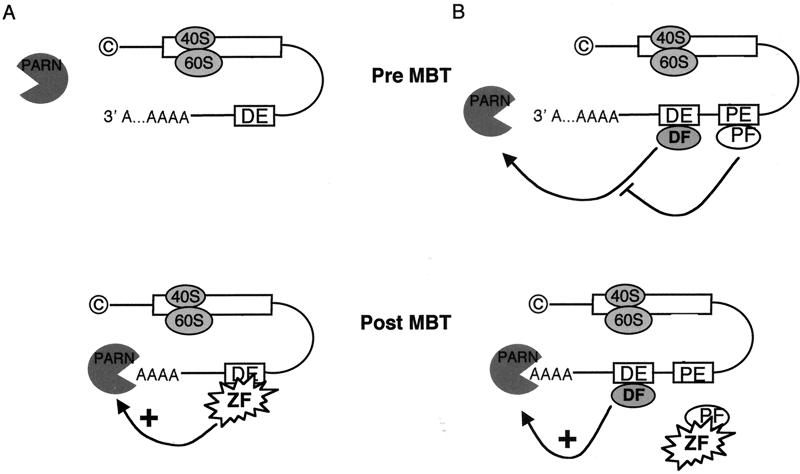
At the midblastula transition, the Xenopus laevis embryonic cell cycle is remodeled from rapid alternations between S and M phases to become the complex adult cell cycle. Cell cycle remodeling occurs after zygotic transcription initiates and is accompanied by terminal downregulation of maternal cyclins A1 and B2. We report here that the disappearance of both cyclin A1 and B2 proteins is preceded by the rapid deadenylation of their mRNAs. A specific mechanism triggers this deadenylation. This mechanism depends upon discrete regions of the 3' untranslated regions and requires zygotic transcription. Together, these results strongly suggest that zygote-dependent deadenylation of cyclin A1 and cyclin B2 mRNAs is responsible for the downregulation of these proteins. These studies also raise the possibility that zygotic control of maternal cyclins plays a role in establishing the adult cell cycle.
Figures
Similar articles
-
ElrA and AUF1 differentially bind cyclin B2 mRNA.Biochem Biophys Res Commun. 2008 Dec 12;377(2):653-657. doi: 10.1016/j.bbrc.2008.10.029. Epub 2008 Oct 16. Biochem Biophys Res Commun. 2008. PMID: 18930026 Free PMC article.
-
Zygotic control of maternal cyclin A1 translation and mRNA stability.Dev Dyn. 2002 Dec;225(4):511-21. doi: 10.1002/dvdy.10191. Dev Dyn. 2002. PMID: 12454927
-
Deadenylation of maternal mRNAs mediated by miR-427 in Xenopus laevis embryos.RNA. 2009 Dec;15(12):2351-63. doi: 10.1261/rna.1882009. Epub 2009 Oct 23. RNA. 2009. PMID: 19854872 Free PMC article.
-
The maternal-zygotic transition: death and birth of RNAs.Science. 2007 Apr 20;316(5823):406-7. doi: 10.1126/science.1140693. Science. 2007. PMID: 17446392 Review.
-
Unmasking the role of the 3' UTR in the cytoplasmic polyadenylation and translational regulation of maternal mRNAs.Bioessays. 1994 Aug;16(8):533-5. doi: 10.1002/bies.950160804. Bioessays. 1994. PMID: 8086000 Review.
Cited by
-
Gene expression during the oocyte-to-embryo transition in mammals.Mol Reprod Dev. 2009 Sep;76(9):805-18. doi: 10.1002/mrd.21038. Mol Reprod Dev. 2009. PMID: 19363788 Free PMC article. Review.
-
ElrA and AUF1 differentially bind cyclin B2 mRNA.Biochem Biophys Res Commun. 2008 Dec 12;377(2):653-657. doi: 10.1016/j.bbrc.2008.10.029. Epub 2008 Oct 16. Biochem Biophys Res Commun. 2008. PMID: 18930026 Free PMC article.
-
Developmental series of gene expression clarifies maternal mRNA provisioning and maternal-to-zygotic transition in a reef-building coral.BMC Genomics. 2021 Nov 11;22(1):815. doi: 10.1186/s12864-021-08114-y. BMC Genomics. 2021. PMID: 34763678 Free PMC article.
-
A conserved maternal-specific repressive domain in Zelda revealed by Cas9-mediated mutagenesis in Drosophila melanogaster.PLoS Genet. 2017 Dec 19;13(12):e1007120. doi: 10.1371/journal.pgen.1007120. eCollection 2017 Dec. PLoS Genet. 2017. PMID: 29261646 Free PMC article.
-
Recycling of Uridylated mRNAs in Starfish Embryos.Biomolecules. 2024 Dec 16;14(12):1610. doi: 10.3390/biom14121610. Biomolecules. 2024. PMID: 39766317 Free PMC article.
References
-
- Barnhart K M. Simplified PCR-mediated, linker-scanning mutagenesis. BioTechniques. 1999;26:624–626. - PubMed
-
- Brewer G. Characterization of c-myc 3′ to 5′ mRNA decay activities in an in vitro system. J Biol Chem. 1998;273:34770–34774. - PubMed
-
- Chevalier S, Couturier A, Chartrain I, Le Guellec R, Beckhelling C, Le Guellec K, Philippe M, Ford C C. Xenopus cyclin E, a nuclear phosphoprotein, accumulates when oocytes gain the ability to initiate DNA replication. J Cell Sci. 1996;109:1173–1184. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources