

An antiprion effect of the anticytoskeletal drug latrunculin A in yeast
- PMID: 11243411
- PMCID: PMC5964936
- DOI: 10.3727/000000001783992650
An antiprion effect of the anticytoskeletal drug latrunculin A in yeast
Abstract
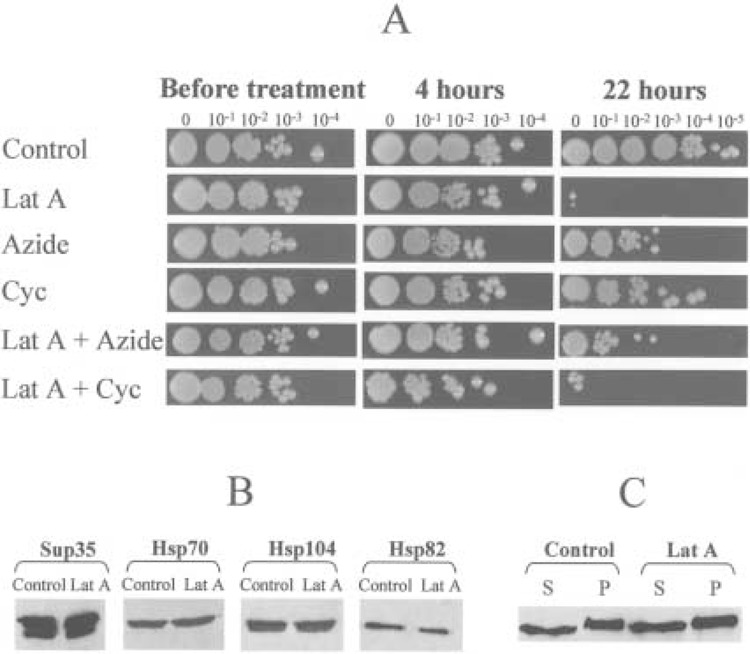
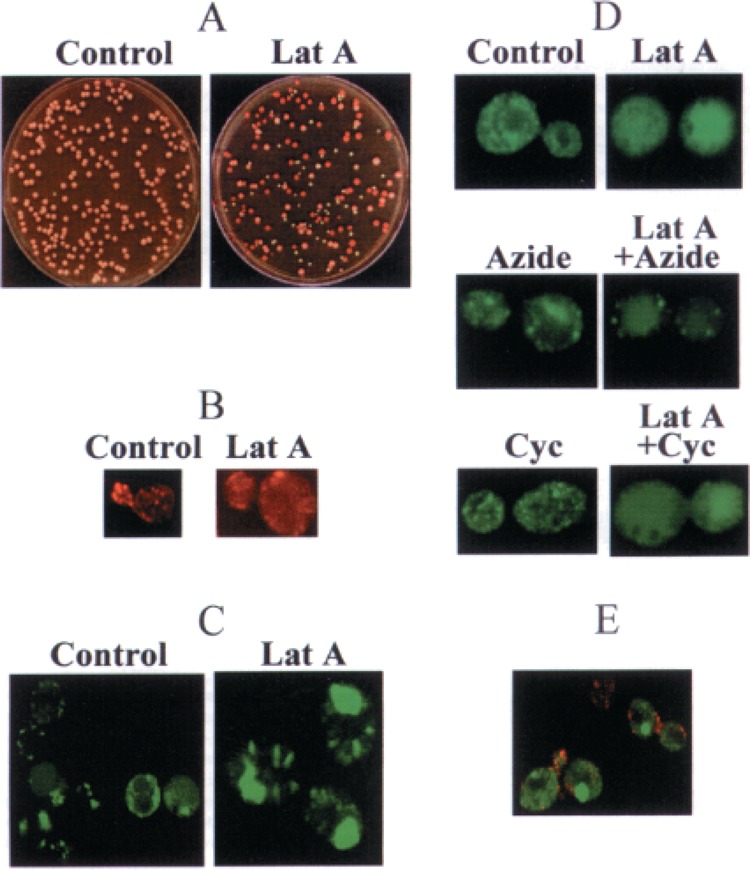
Prions are infectious aggregation-prone isoforms of the normal proteins, supposedly able to seed aggregation of the normal cellular counterparts. In vitro, prion proteins form amyloid fibers, resembling cytoskeletal structures. Yeast prion [PSI], which is a cytoplasmically inherited aggregated isoform of the translation termination factor Sup35p (eRF3), serves as a useful model for studying mechanisms of prion diseases and other amyloidoses. The previously described interaction between Sup35p and cytoskeletal assembly protein Sla1p points to the possible relationships between prions and cytoskeletal networks. Although the Sup35PSI+ aggregates do not colocalize with actin patches, we have shown that yeast cells are efficiently cured of the [PSI] prion by prolonged incubation with latrunculin A, a drug disrupting the actin cytoskeleton. On the other hand, treatments with sodium azide or cycloheximide, agents blocking yeast protein synthesis and cell proliferation but not disrupting the cytoskeleton, do not cause a significant loss of [PSI]. Moreover, simultaneous treatment with sodium azide or cycloheximide blocks [PSI] curing by latrunculin A, indicating that prion loss in the presence of latrunculin A requires a continuation of protein synthesis during cytoskeleton disruption. The sodium azide treatment also decreases the toxic effect of latrunculin A. Latrunculin A influences neither the levels of total cellular Sup35p nor the levels of chaperone proteins, such as Hsp104 and Hsp70, which were previously shown to affect [PSI]. This makes an indirect effect of latrunculin A on [PSI] via induction of Hsps unlikely. Fluorescence microscopy detects changes in the structure and/or localization of the Sup35PSI+ aggregates in latrunculin A-treated cells. We conclude that the stable maintenance of the [PSI] prion aggregates in the protein-synthesizing yeast cells partly depends on an intact actin cytoskeleton, suggesting that anticytoskeletal treatments could be used to counteract some aggregation-related disorders.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases