

Estrogen biphasically modifies hypothalamic GABAergic function concomitantly with negative and positive control of luteinizing hormone release
- PMID: 11245692
- PMCID: PMC6762630
- DOI: 10.1523/JNEUROSCI.21-06-02085.2001
Estrogen biphasically modifies hypothalamic GABAergic function concomitantly with negative and positive control of luteinizing hormone release
Abstract
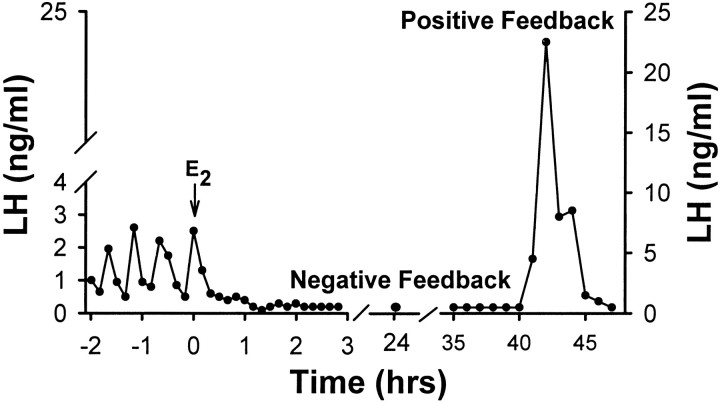
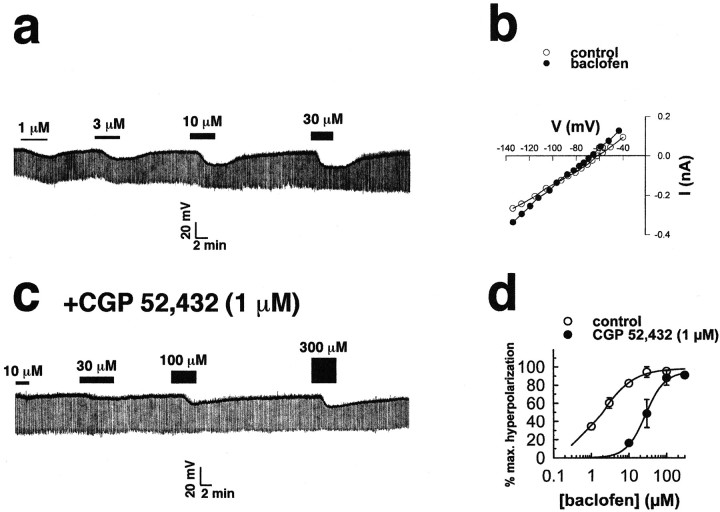
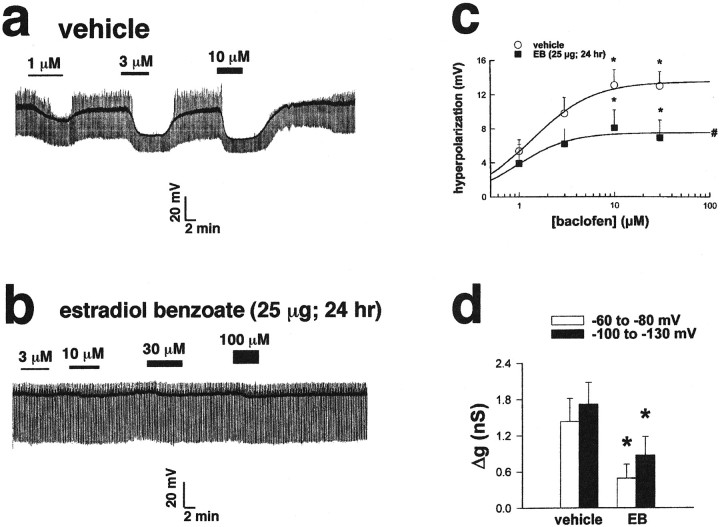
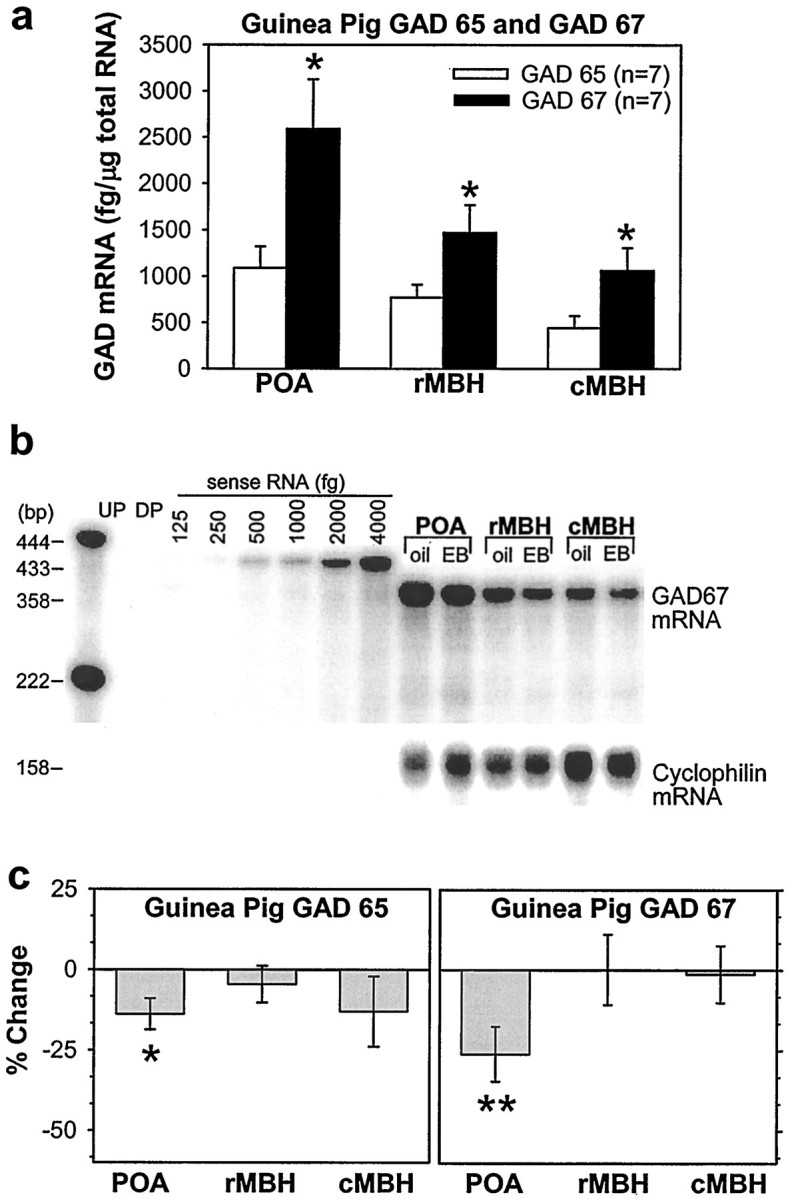
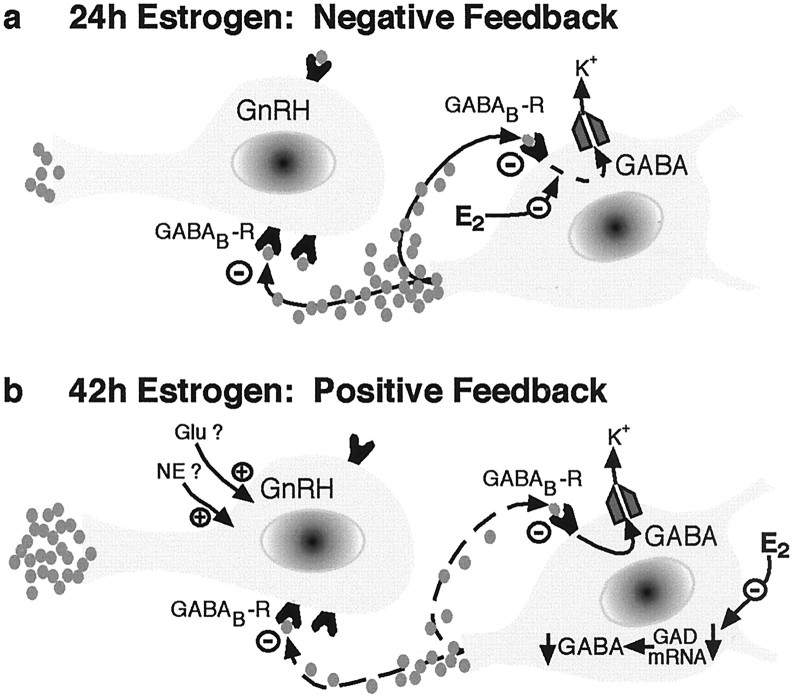
The principal role of estrogen is its control of the female ovulatory cycle via negative and positive feedback on gonadotropin secretion. However, a detailed, cohesive picture of how the steroid specifically regulates the excitability of hypothalamic neurons involved in the central control of gonadotropin secretion is still emerging. Here, we used an ovariectomized female guinea pig model to test the hypothesis that estrogen acts on GABAergic neurons in the preoptic area (POA) to elicit a biphasic profile of luteinizing hormone (LH) secretion. Intracellular electrophysiological recordings revealed that estradiol benzoate (EB; 25 microgram, s.c.) decreased the hyperpolarizing response of GABAergic neurons to the GABA(B) receptor agonist baclofen 24 hr after treatment. This effect of GABA(B) receptor stimulation in unidentified POA neurons was still depressed 42 hr after EB administration. By the use of a ribonuclease protection assay, however, EB reduced glutamic acid decarboxylase mRNA expression 42 hr but not 24 hr after its administration. Thus, estrogen attenuated the autoinhibition of GABAergic POA neurons during the initial LH suppressive (i.e., negative feedback) phase and subsequently reduced GABAergic function during the LH surge (i.e., positive feedback). These studies demonstrate that the effects of estrogen on hypothalamic GABAergic neurons coincide with the inhibitory and stimulatory actions, respectively, of the steroid on LH secretion. Furthermore, the data provide novel insights into the mechanism by which estrogen regulates hypothalamic GABAergic neurons, which are critical for the biphasic modulation of LH release observed over the course of the female ovulatory cycle.
Figures

References
-
- Becker JB, Snyder PJ, Miller MM, Westgate SA, Jenuwine MJ. The influence of the estrous cycle and intrastriatal estradiol on sensorimotor performance in the female rat. Pharmacol Biochem Behav. 1987;27:53–59. - PubMed
-
- Bowery N. GABA(B) receptors and their significance in mammalian pharmacology. Trends Pharmacol Sci. 1989;10:401–407. - PubMed
-
- Brann DW, Zamorano PL, Putnam-Roberts CD, Mahesh VB. Gamma-aminobutyric acid-opioid interactions in the regulation of gonadotropin secretion in the immature female rat. Neuroendocrinology. 1992;56:445–452. - PubMed
-
- Brown D, Herbison AE, Robinson JE, Marrs RW, Leng G. Modelling the luteinizing hormone-releasing hormone pulse generator. Neuroscience. 1994;63:869–879. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources