

Symmetry, stereotypy, and topography of odorant representations in mouse olfactory bulbs
- PMID: 11245695
- PMCID: PMC6762614
- DOI: 10.1523/JNEUROSCI.21-06-02113.2001
Symmetry, stereotypy, and topography of odorant representations in mouse olfactory bulbs
Abstract
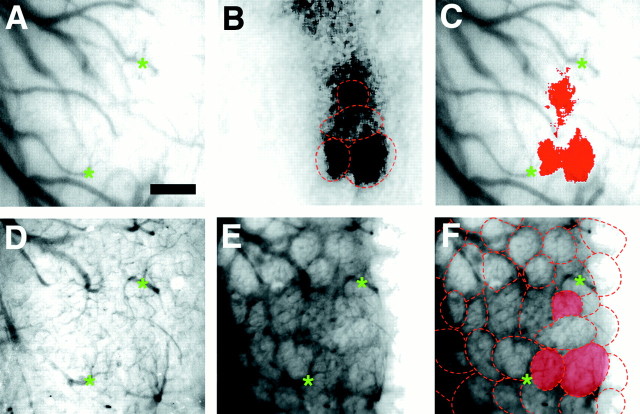
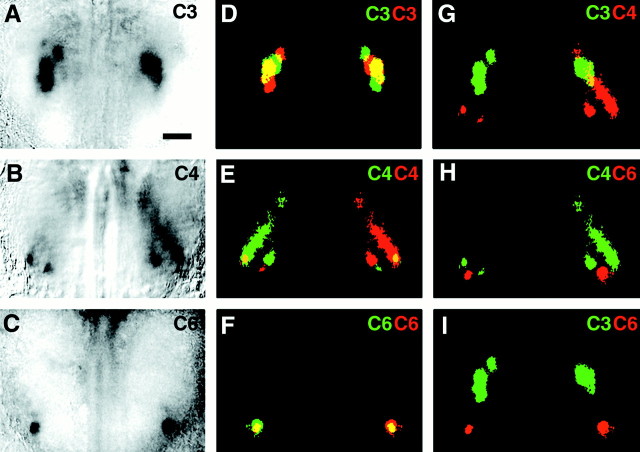
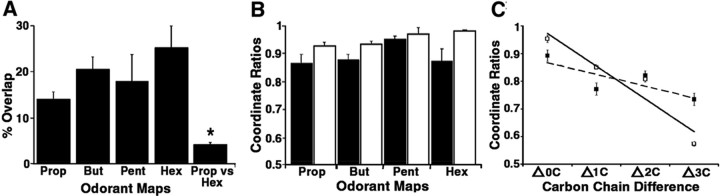
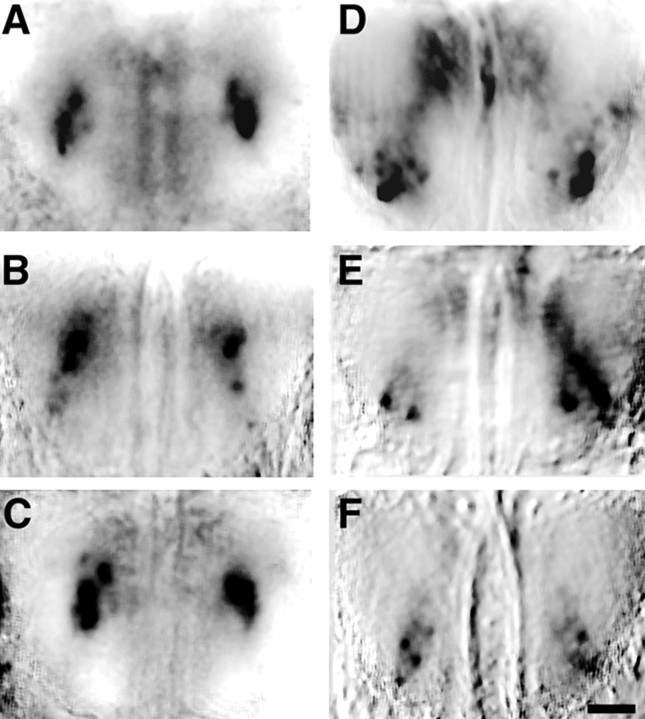
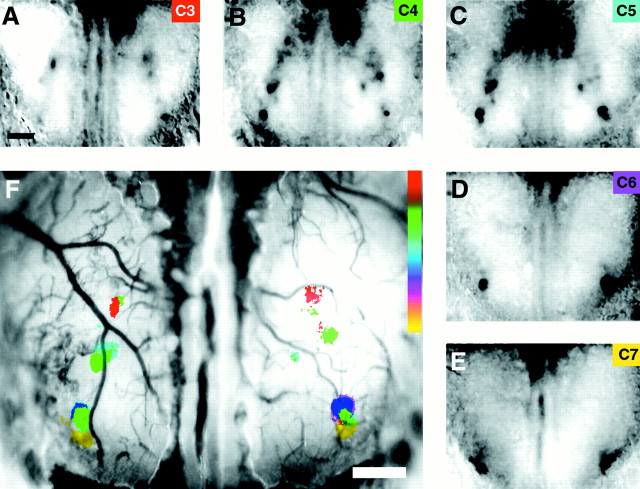
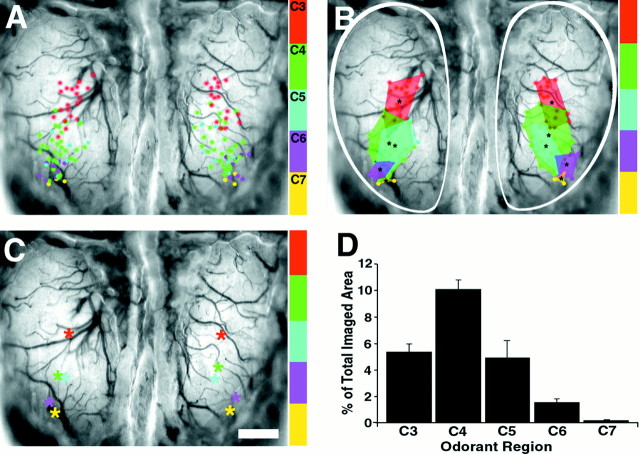
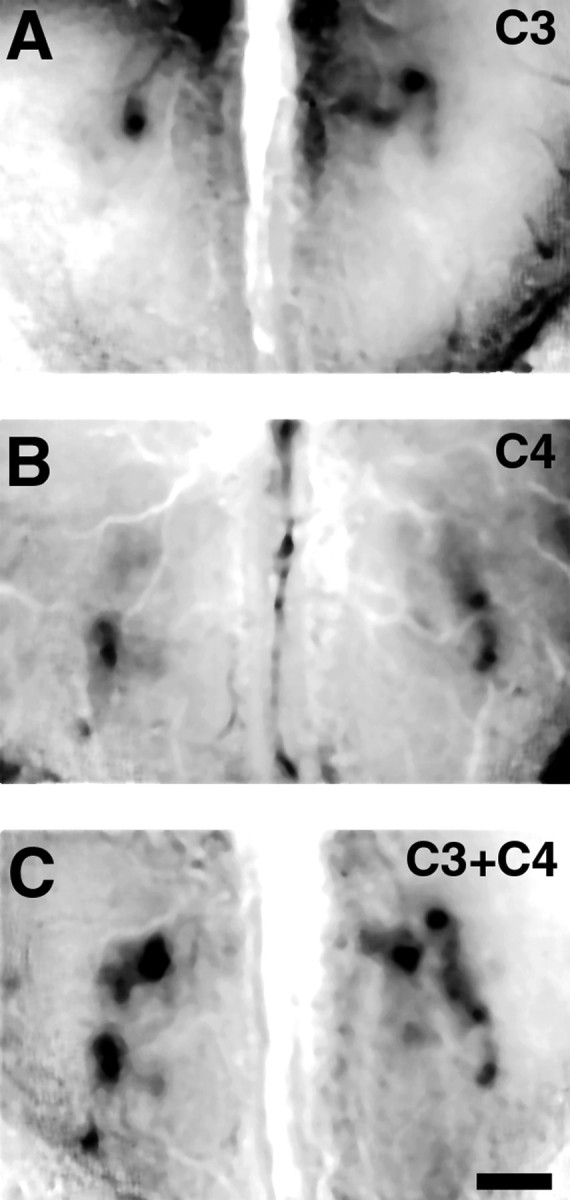
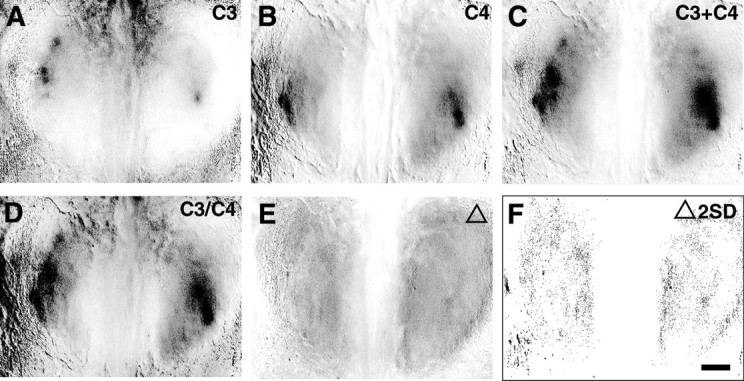
The molecular basis of vertebrate odorant representations has been derived extensively from mice. The functional correlates of these molecular features were visualized using optical imaging of intrinsic signals in mouse olfactory bulbs. Single odorants activated clusters of glomeruli in consistent, restricted portions of the bulb. Patterns of activated glomeruli were clearly bilaterally symmetric and consistent in different individual mice, but the precise number, position, and intensity of activated glomeruli in the two bulbs of the same individual and between individuals varied considerably. Representations of aliphatic aldehydes of different carbon chain length shifted systematically along a rostral-caudal strip of the dorsal bulb, indicating a functional topography of odorant representations. Binary mixtures of individual aldehydes elicited patterns of glomerular activation that were topographic combinations of the maps for each individual odor. Thus the principles derived from the molecular organization of a small subset of murine olfactory receptor neuron projection patterns-bilateral symmetry, local clustering, and local variability-are reliable guides to the initial functional representation of odorant molecules.
Figures
References
-
- Brunjes PC. Unilateral naris closure and olfactory system development. Brain Res Brain Res Rev. 1994;19:146–160. - PubMed
-
- Cain WS, de Wijk R, Lulejian C, Schiet F, See LC. Odor identification: perceptual and semantic dimensions. Chem Senses. 1998;23:309–326. - PubMed
-
- Christensen TA, Pawlowski VM, Lei H, Hildebrand JG. Multi-unit recordings reveal context-dependent modulation of synchrony in odor-specific neural ensembles. Nat Neurosci. 2000;3:927–931. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources