

Herpes simplex virus DNA packaging sequences adopt novel structures that are specifically recognized by a component of the cleavage and packaging machinery
- PMID: 11248036
- PMCID: PMC30611
- DOI: 10.1073/pnas.061555698
Herpes simplex virus DNA packaging sequences adopt novel structures that are specifically recognized by a component of the cleavage and packaging machinery
Abstract
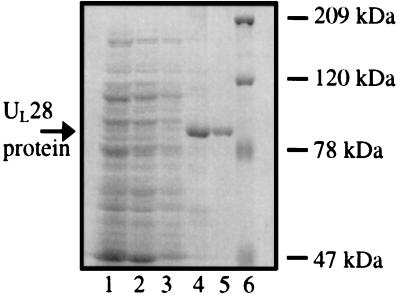
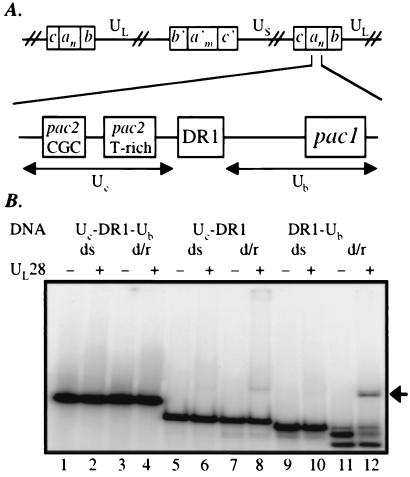
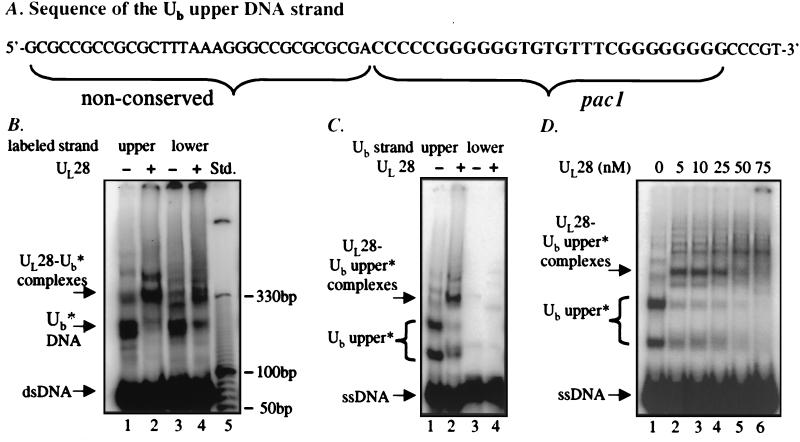
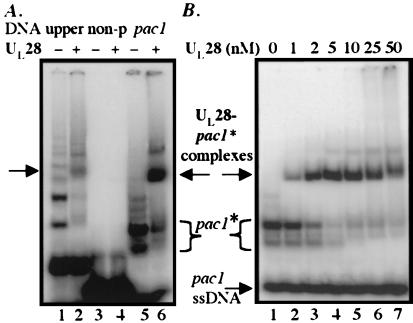
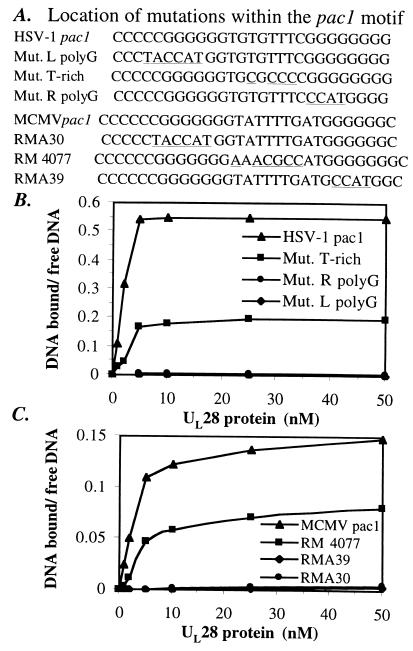
The product of the herpes simplex virus type 1 U(L)28 gene is essential for cleavage of concatemeric viral DNA into genome-length units and packaging of this DNA into viral procapsids. To address the role of U(L)28 in this process, purified U(L)28 protein was assayed for the ability to recognize conserved herpesvirus DNA packaging sequences. We report that DNA fragments containing the pac1 DNA packaging motif can be induced by heat treatment to adopt novel DNA conformations that migrate faster than the corresponding duplex in nondenaturing gels. Surprisingly, these novel DNA structures are high-affinity substrates for U(L)28 protein binding, whereas double-stranded DNA of identical sequence composition is not recognized by U(L)28 protein. We demonstrate that only one strand of the pac1 motif is responsible for the formation of novel DNA structures that are bound tightly and specifically by U(L)28 protein. To determine the relevance of the observed U(L)28 protein-pac1 interaction to the cleavage and packaging process, we have analyzed the binding affinity of U(L)28 protein for pac1 mutants previously shown to be deficient in cleavage and packaging in vivo. Each of the pac1 mutants exhibited a decrease in DNA binding by U(L)28 protein that correlated directly with the reported reduction in cleavage and packaging efficiency, thereby supporting a role for the U(L)28 protein-pac1 interaction in vivo. These data therefore suggest that the formation of novel DNA structures by the pac1 motif confers added specificity on recognition of DNA packaging sequences by the U(L)28-encoded component of the herpesvirus cleavage and packaging machinery.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
