

Structure, folding and activity of the VS ribozyme: importance of the 2-3-6 helical junction
- PMID: 11250907
- PMCID: PMC145516
- DOI: 10.1093/emboj/20.6.1415
Structure, folding and activity of the VS ribozyme: importance of the 2-3-6 helical junction
Abstract
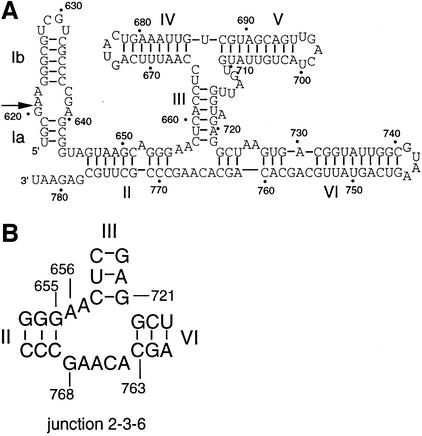
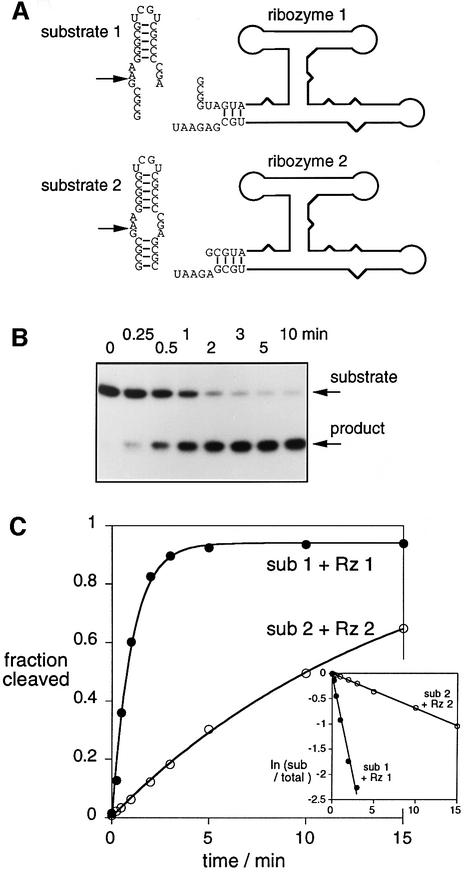
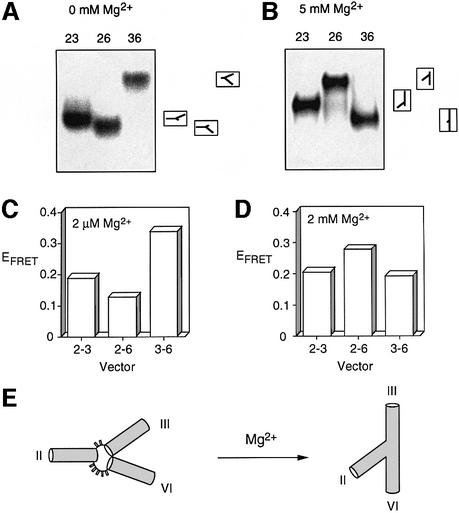
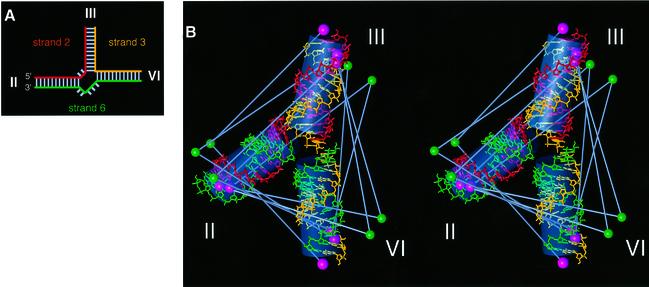
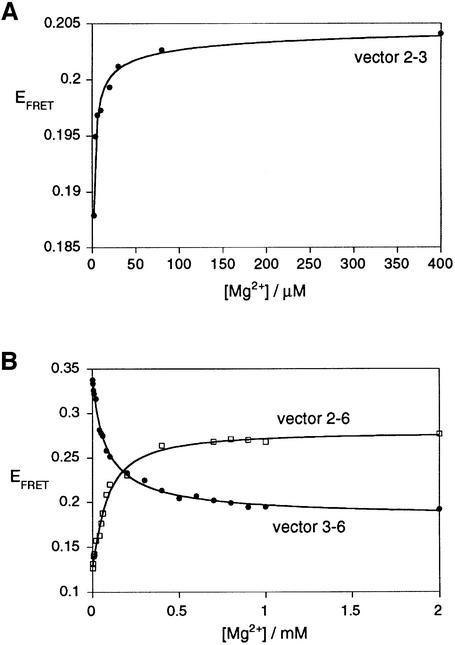
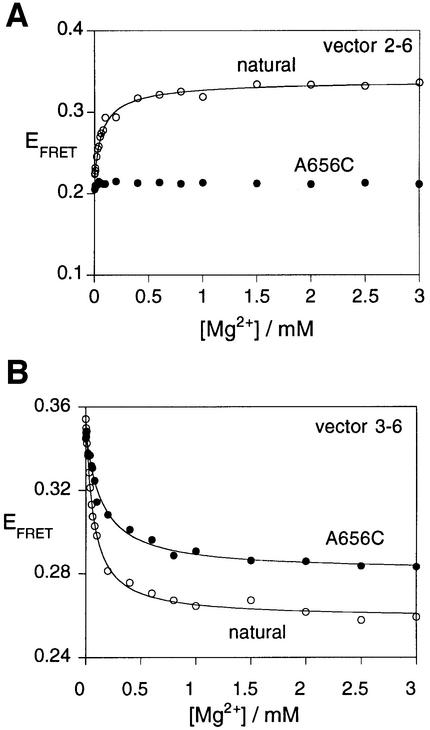
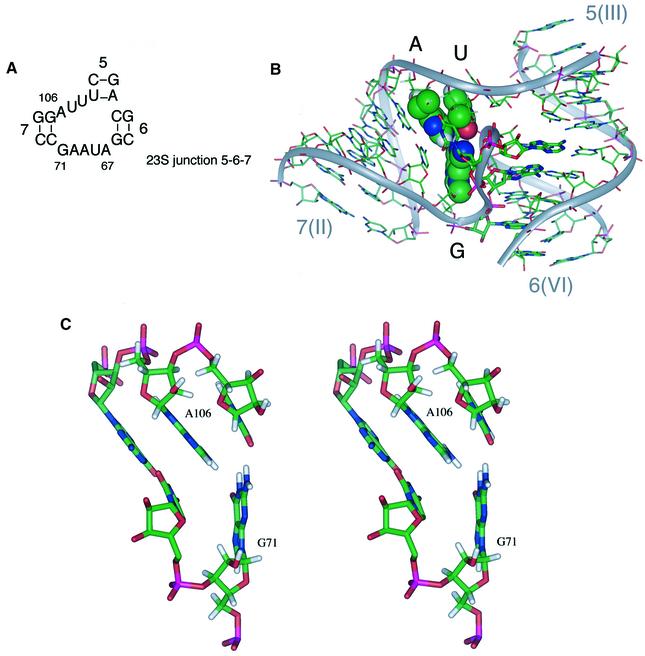
The VS nucleolytic ribozyme has a core comprising five helices organized by two three-way junctions. The ribozyme can act in trans on a hairpin-loop substrate, with which it interacts via tertiary contacts. We have determined that one of the junctions (2-3-6) undergoes two-stage ion-dependent folding into a stable conformation, and have determined the global structure of the folded junction using long-range distance restraints derived from fluorescence resonance energy transfer. A number of sequence variants in the junction are severely impaired in ribozyme cleavage, and there is good correlation between changes in activity and alteration in the folding of junction 2-3-6. These studies point to a special importance of G and A nucleotides immediately adjacent to helix II, and comparison with a similar junction of known structure indicates that this could adopt a guanine-wedge structure. We propose that the 2-3-6 junction organizes important aspects of the structure of the ribozyme to facilitate productive association with the substrate, and suggest that this results in an interaction between the substrate and the A730 loop to create the active complex.
Figures
Similar articles
-
Folding and catalysis by the VS ribozyme.Biochimie. 2002 Sep;84(9):889-96. doi: 10.1016/s0300-9084(02)01406-2. Biochimie. 2002. PMID: 12458081
-
The structure and active site of the Varkud satellite ribozyme.Biochem Soc Trans. 2002 Nov;30(Pt 6):1170-5. doi: 10.1042/bst0301170. Biochem Soc Trans. 2002. PMID: 12440998 Review.
-
Structure and activity of the hairpin ribozyme in its natural junction conformation: effect of metal ions.Biochemistry. 1998 Oct 6;37(40):14195-203. doi: 10.1021/bi981513+. Biochemistry. 1998. PMID: 9760257
-
NMR structure of the A730 loop of the Neurospora VS ribozyme: insights into the formation of the active site.Nucleic Acids Res. 2011 May;39(10):4427-37. doi: 10.1093/nar/gkq1244. Epub 2011 Jan 25. Nucleic Acids Res. 2011. PMID: 21266483 Free PMC article.
-
The Varkud satellite ribozyme.RNA. 2004 Feb;10(2):151-8. doi: 10.1261/rna.5217104. RNA. 2004. PMID: 14730013 Free PMC article. Review.
Cited by
-
Constitutive regulatory activity of an evolutionarily excluded riboswitch variant.J Biol Chem. 2011 Aug 5;286(31):27406-15. doi: 10.1074/jbc.M111.229047. Epub 2011 Jun 15. J Biol Chem. 2011. PMID: 21676871 Free PMC article.
-
Crystal structure of the Varkud satellite ribozyme.Nat Chem Biol. 2015 Nov;11(11):840-6. doi: 10.1038/nchembio.1929. Epub 2015 Sep 28. Nat Chem Biol. 2015. PMID: 26414446 Free PMC article.
-
Rapid formation of a solvent-inaccessible core in the Neurospora Varkud satellite ribozyme.EMBO J. 2001 Oct 1;20(19):5461-9. doi: 10.1093/emboj/20.19.5461. EMBO J. 2001. PMID: 11574478 Free PMC article.
-
The global structure of the VS ribozyme.EMBO J. 2002 May 15;21(10):2461-71. doi: 10.1093/emboj/21.10.2461. EMBO J. 2002. PMID: 12006498 Free PMC article.
-
Mechanisms of RNA catalysis.Philos Trans R Soc Lond B Biol Sci. 2011 Oct 27;366(1580):2910-7. doi: 10.1098/rstb.2011.0132. Philos Trans R Soc Lond B Biol Sci. 2011. PMID: 21930582 Free PMC article.
References
-
- Andersen A.A. and Collins,R.A. (2000) Rearrangement of a stable RNA secondary structure during VS ribozyme catalysis. Mol. Cell, 5, 469–478. - PubMed
-
- Ban N., Nissen,P., Hansen,J., Moore,P.B. and Steitz,T.A. (2000) The complete atomic structure of the large ribosomal subunit at 2.4 Å resolution. Science, 289, 905–920. - PubMed
-
- Bassi G., Møllegaard,N.E., Murchie,A.I.H., von Kitzing,E. and Lilley,D.M.J. (1995) Ionic interactions and the global conformations of the hammerhead ribozyme. Nature Struct. Biol., 2, 45–55. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
