

CCR6-deficient mice have impaired leukocyte homeostasis and altered contact hypersensitivity and delayed-type hypersensitivity responses
- PMID: 11254677
- PMCID: PMC208945
- DOI: 10.1172/JCI11297
CCR6-deficient mice have impaired leukocyte homeostasis and altered contact hypersensitivity and delayed-type hypersensitivity responses
Abstract
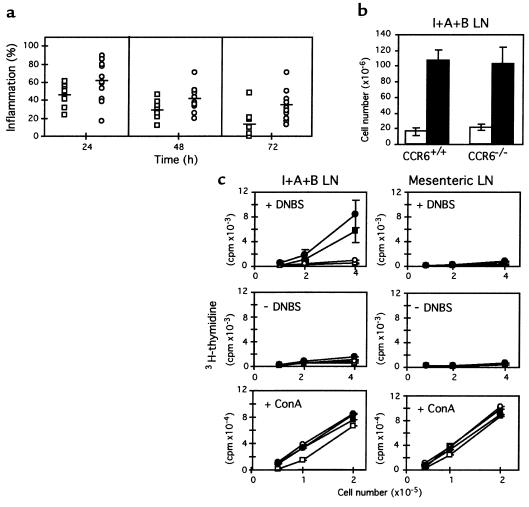
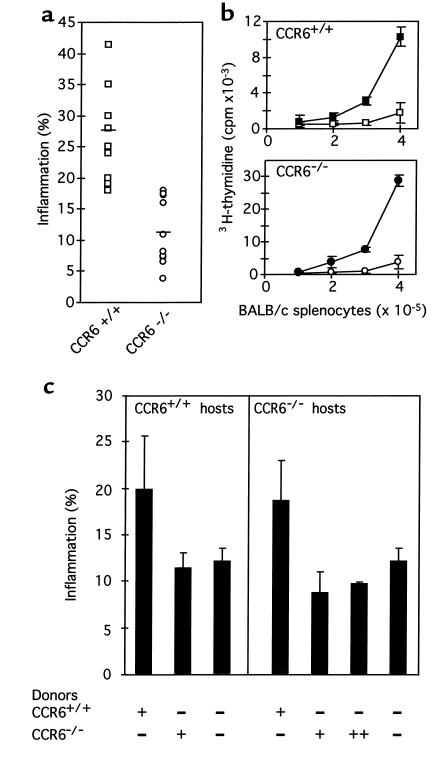
CCR6 expression in dendritic, T, and B cells suggests that this beta-chemokine receptor may regulate the migration and recruitment of antigen-presenting and immunocompetent cells during inflammatory and immunological responses. Here we demonstrate that CCR6-/- mice have underdeveloped Peyer's patches, in which the myeloid CD11b+ CD11c+ dendritic-cell subset is not present in the subepithelial dome. CCR6-/- mice also have increased numbers in T-cell subpopulations within the intestinal mucosa. In 2,4-dinitrofluorobenzene-induced contact hypersensitivity (CHS) studies, CCR6-/- mice developed more severe and more persistent inflammation than wild-type (WT) animals. Conversely, in a delayed-type hypersensitivity (DTH) model induced with allogeneic splenocytes, CCR6-/- mice developed no inflammatory response. The altered responses seen in the CHS and DTH assays suggest the existence of a defect in the activation and/or migration of the CD4(+) T-cell subsets that downregulate or elicit the inflammation response, respectively. These findings underscore the role of CCR6 in cutaneous and intestinal immunity and the utility of CCR6-/- mice as a model to study pathologies in these tissues. This article was published online in advance of the print edition. The date of publication is available from the JCI website, http://www.jci.org.
Figures
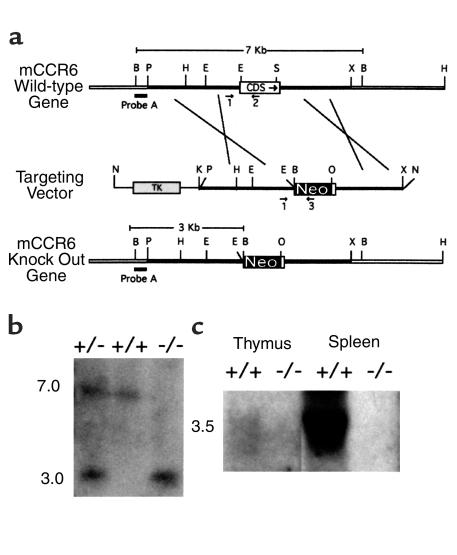
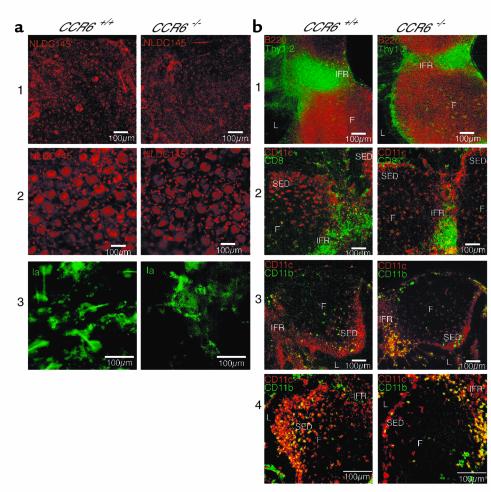
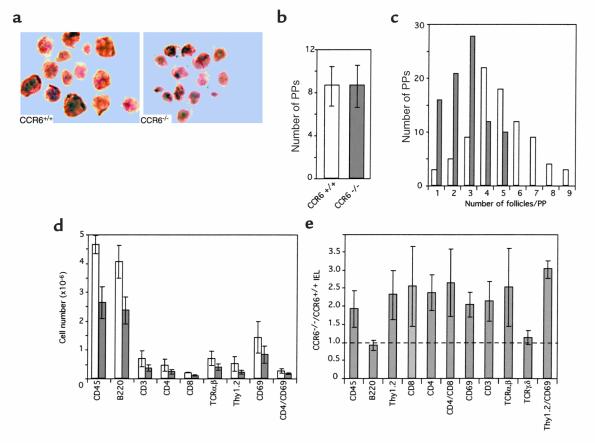
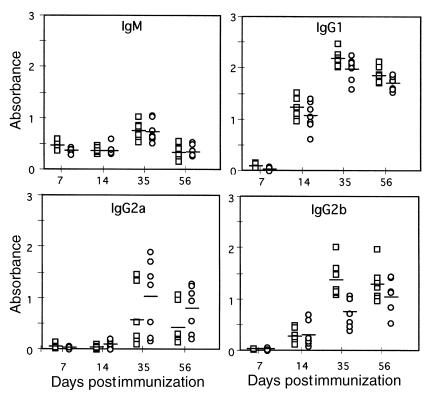
References
-
- Banchereau J, et al. Immunobiology of dendritic cells. Annu Rev Immunol. 2000;18:767–811. - PubMed
-
- Forster R, et al. A putative chemokine receptor, BLR1, directs B cell migration to defined lymphoid organs and specific anatomic compartments of the spleen. Cell. 1996;87:1037–1047. - PubMed
-
- Sallusto F, Lanzavecchia A, Mackay CR. Chemokines and chemokine receptors in T-cell priming and Th1/Th2-mediated responses. Immunol Today. 1998;19:568–574. - PubMed
-
- Sallusto F, Lenig D, Forster R, Lipp M, Lanzavecchia A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature. 1999;401:708–712. - PubMed
-
- Forster R, et al. CCR7 coordinates the primary immune response by establishing functional microenvironments in secondary lymphoid organs. Cell. 1999;99:23–33. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
