

UGO1 encodes an outer membrane protein required for mitochondrial fusion
- PMID: 11257114
- PMCID: PMC2199209
- DOI: 10.1083/jcb.152.6.1123
UGO1 encodes an outer membrane protein required for mitochondrial fusion
Erratum in
- J Cell Biol 2001 Apr 30;153(3):635
Abstract
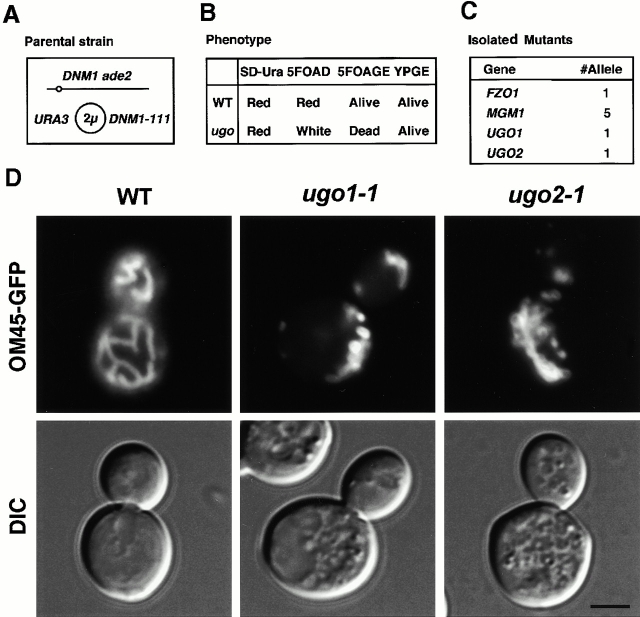
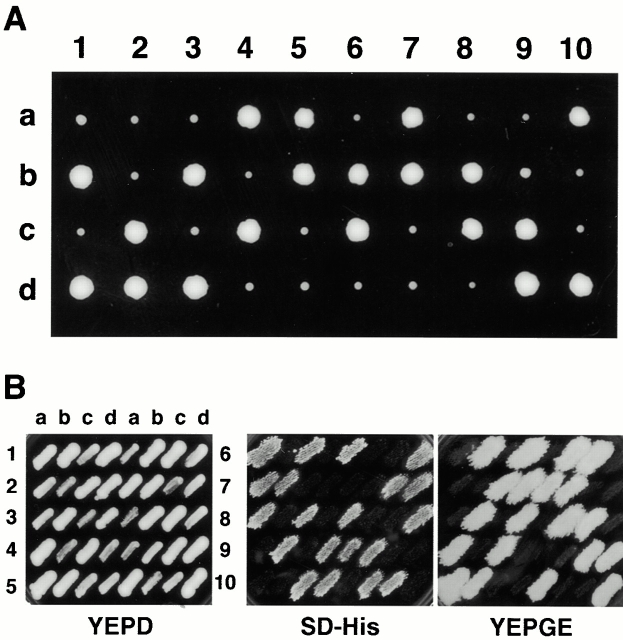
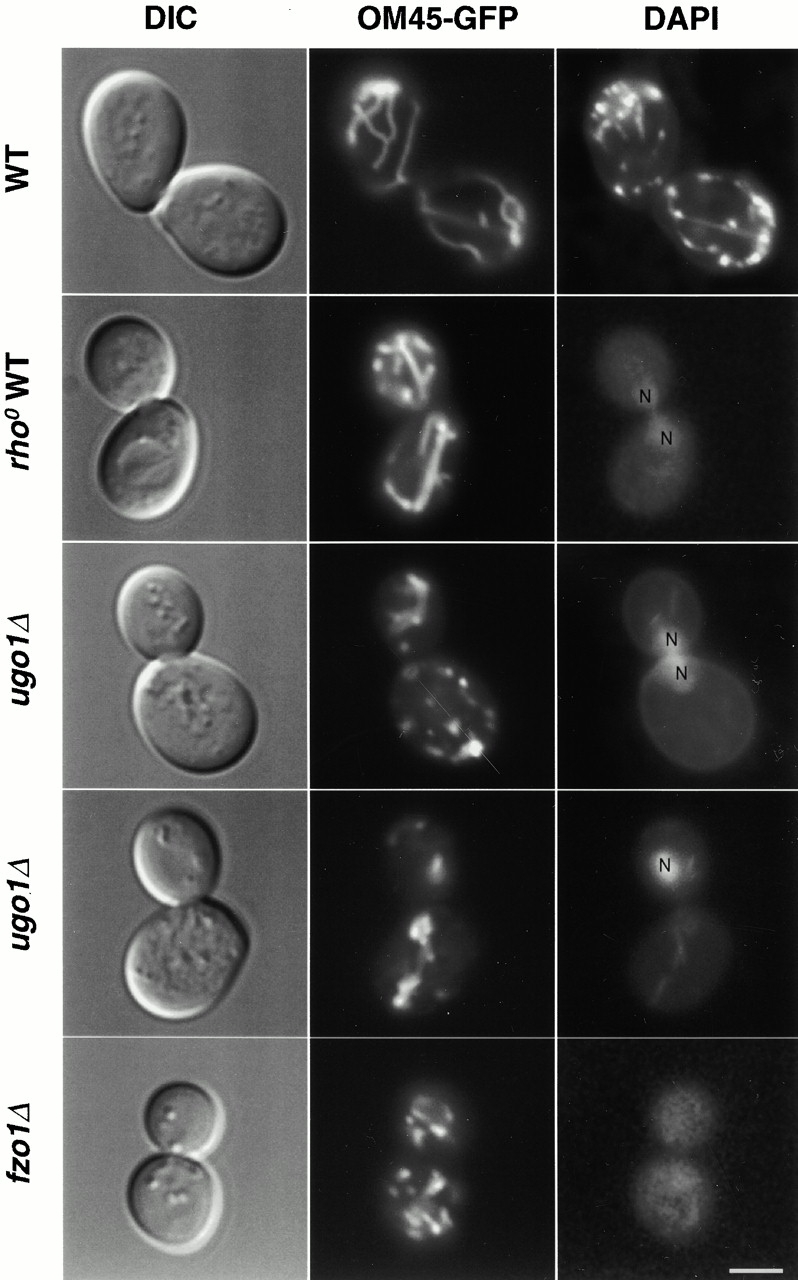
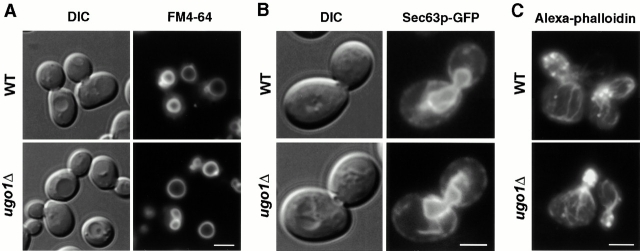
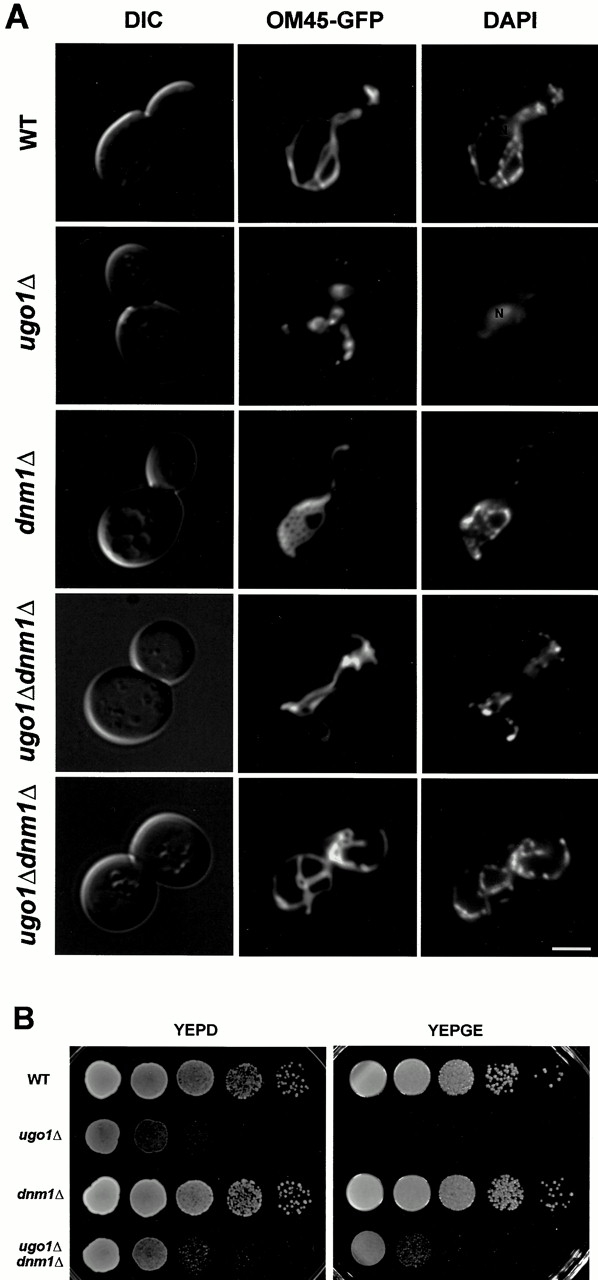
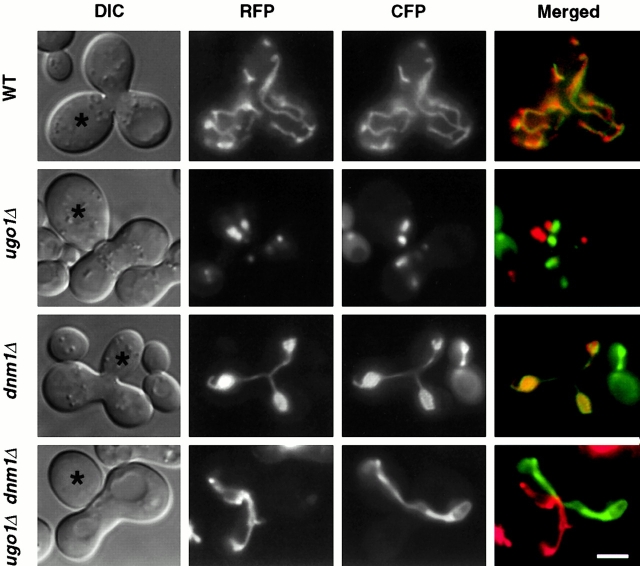
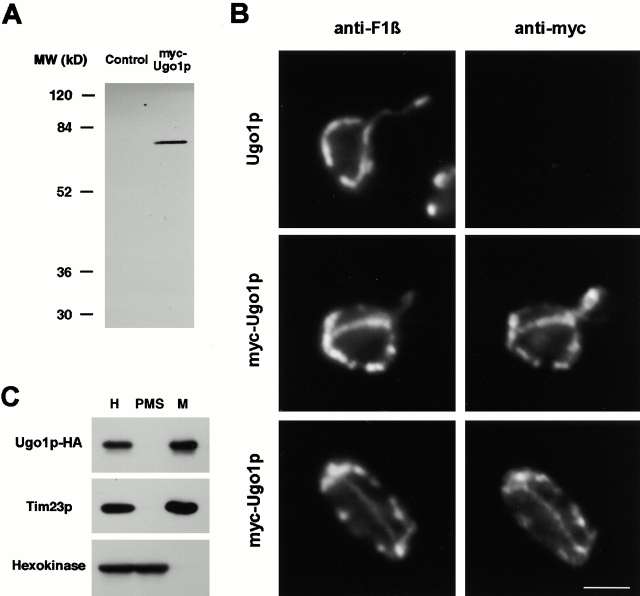
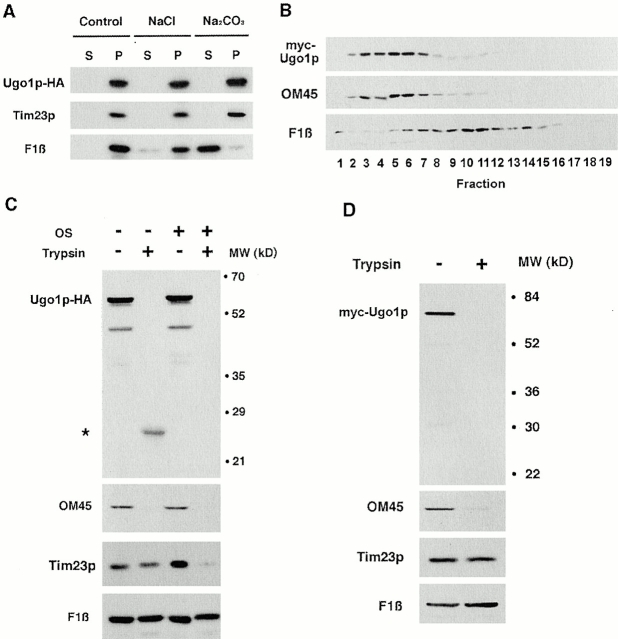
Membrane fusion plays an important role in controlling the shape, number, and distribution of mitochondria. In the yeast Saccharomyces cerevisiae, the outer membrane protein Fzo1p has been shown to mediate mitochondrial fusion. Using a novel genetic screen, we have isolated new mutants defective in the fusion of their mitochondria. One of these mutants, ugo1, shows several similarities to fzo1 mutants. ugo1 cells contain numerous mitochondrial fragments instead of the few long, tubular organelles seen in wild-type cells. ugo1 mutants lose mitochondrial DNA (mtDNA). In zygotes formed by mating two ugo1 cells, mitochondria do not fuse and mix their matrix contents. Fragmentation of mitochondria and loss of mtDNA in ugo1 mutants are rescued by disrupting DNM1, a gene required for mitochondrial division. We find that UGO1 encodes a 58-kD protein located in the mitochondrial outer membrane. Ugo1p appears to contain a single transmembrane segment, with its NH(2) terminus facing the cytosol and its COOH terminus in the intermembrane space. Our results suggest that Ugo1p is a new outer membrane component of the mitochondrial fusion machinery.
Figures
References
-
- Adams A., Gottschling D., Kaiser C., Stearns T. Methods in Yeast Genetics 1997. Cold Spring Harbor Laboratory Press, ; Plainview, NY: pp. 177 pp
-
- Adams A.E., Pringle J.R. Staining of actin with fluorochrome-conjugated phalloidin. Methods Enzymol. 1991;194:729–731. - PubMed
-
- Bereiter-Hahn J., Voth M. Dynamics of mitochondria in living cellsshape changes, dislocations, fusion, and fission of mitochondria. Microsc. Res. Tech. 1994;27:198–219. - PubMed
-
- Boeke J.D., LaCroute F., Fink G.R. A positive selection for mutants lacking orotidine-5′-phosphate decarboxylase activity in yeast5-fluoro-orotic acid resistance. Mol. Gen. Genet. 1984;197:345–346. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
