

Angiomotin: an angiostatin binding protein that regulates endothelial cell migration and tube formation
- PMID: 11257124
- PMCID: PMC2199208
- DOI: 10.1083/jcb.152.6.1247
Angiomotin: an angiostatin binding protein that regulates endothelial cell migration and tube formation
Abstract
Angiostatin, a circulating inhibitor of angiogenesis, was identified by its ability to maintain dormancy of established metastases in vivo. In vitro, angiostatin inhibits endothelial cell migration, proliferation, and tube formation, and induces apoptosis in a cell type-specific manner. We have used a construct encoding the kringle domains 1--4 of angiostatin to screen a placenta yeast two-hybrid cDNA library for angiostatin-binding peptides. Here we report the identification of angiomotin, a novel protein that mediates angiostatin inhibition of migration and tube formation of endothelial cells. In vivo, angiomotin is expressed in the endothelial cells of capillaries as well as larger vessels of the human placenta. Upon expression of angiomotin in HeLa cells, angiomotin bound and internalized fluorescein-labeled angiostatin. Transfected angiomotin as well as endogenous angiomotin protein were localized to the leading edge of migrating endothelial cells. Expression of angiomotin in endothelial cells resulted in increased cell migration, suggesting a stimulatory role of angiomotin in cell motility. However, treatment with angiostatin inhibited migration and tube formation in angiomotin-expressing cells but not in control cells. These findings indicate that angiostatin inhibits cell migration by interfering with angiomotin activity in endothelial cells.
Figures
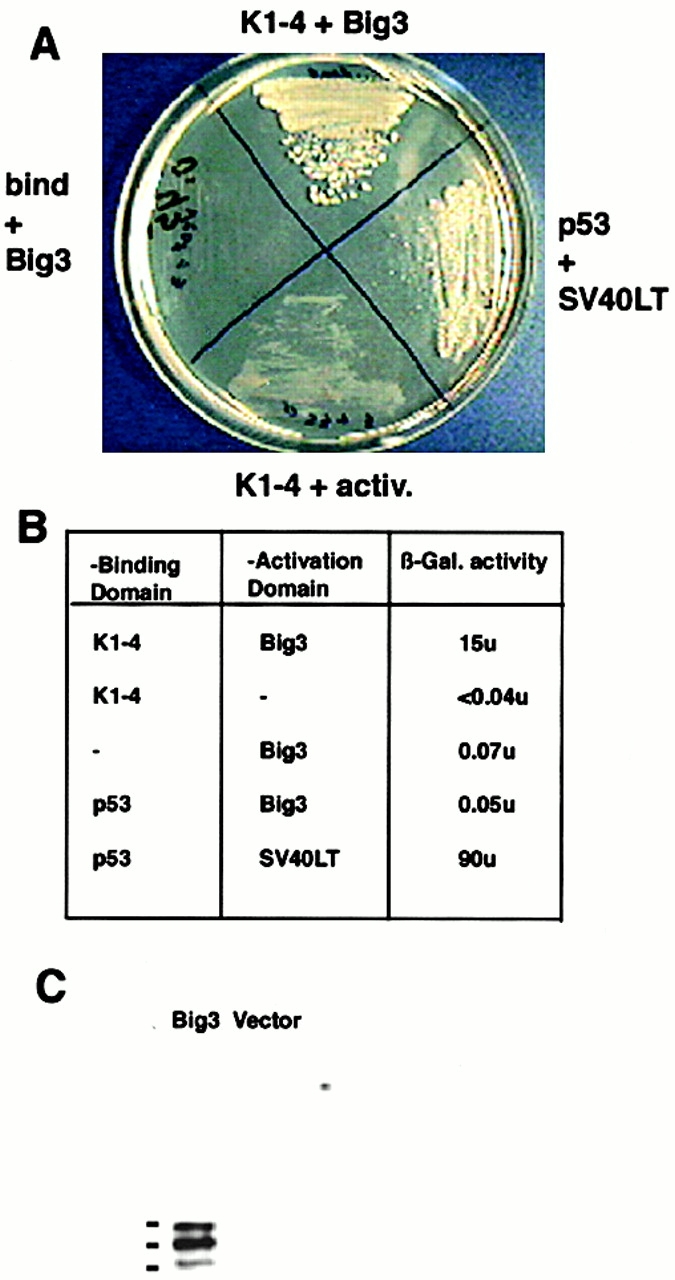
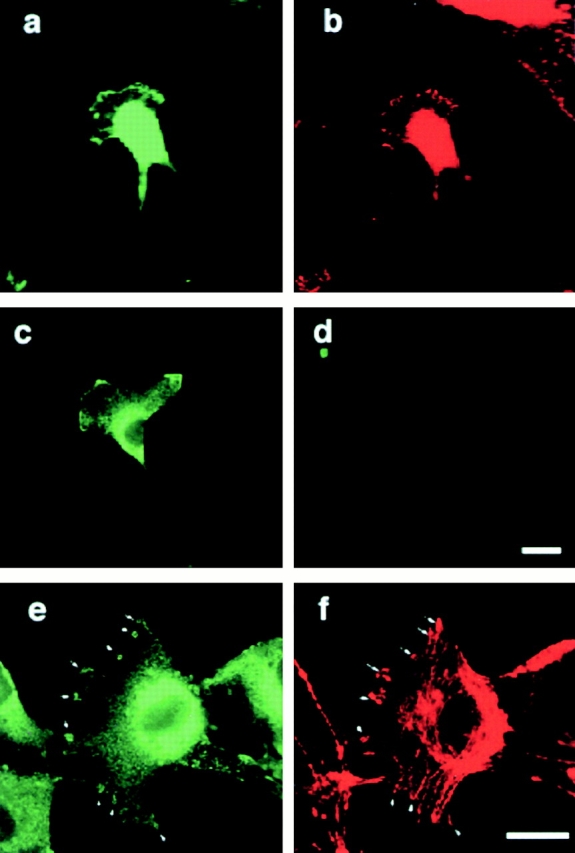
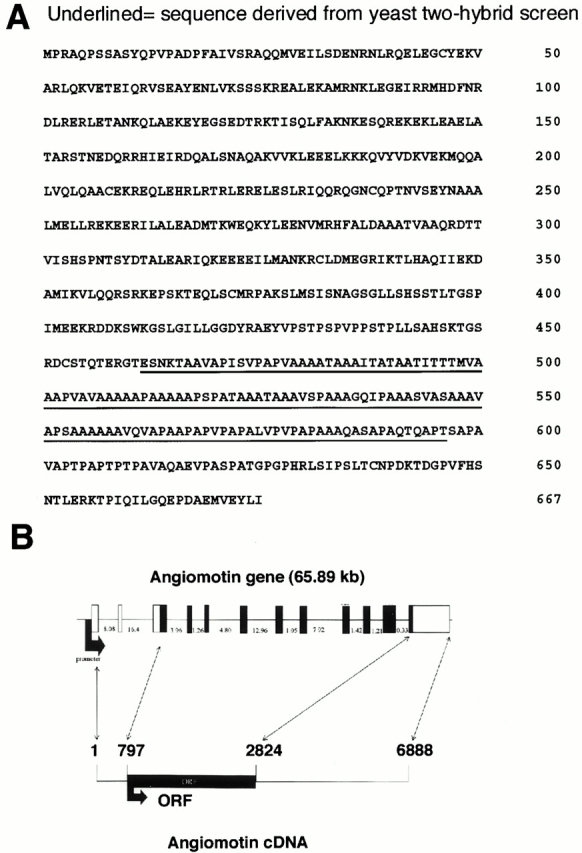
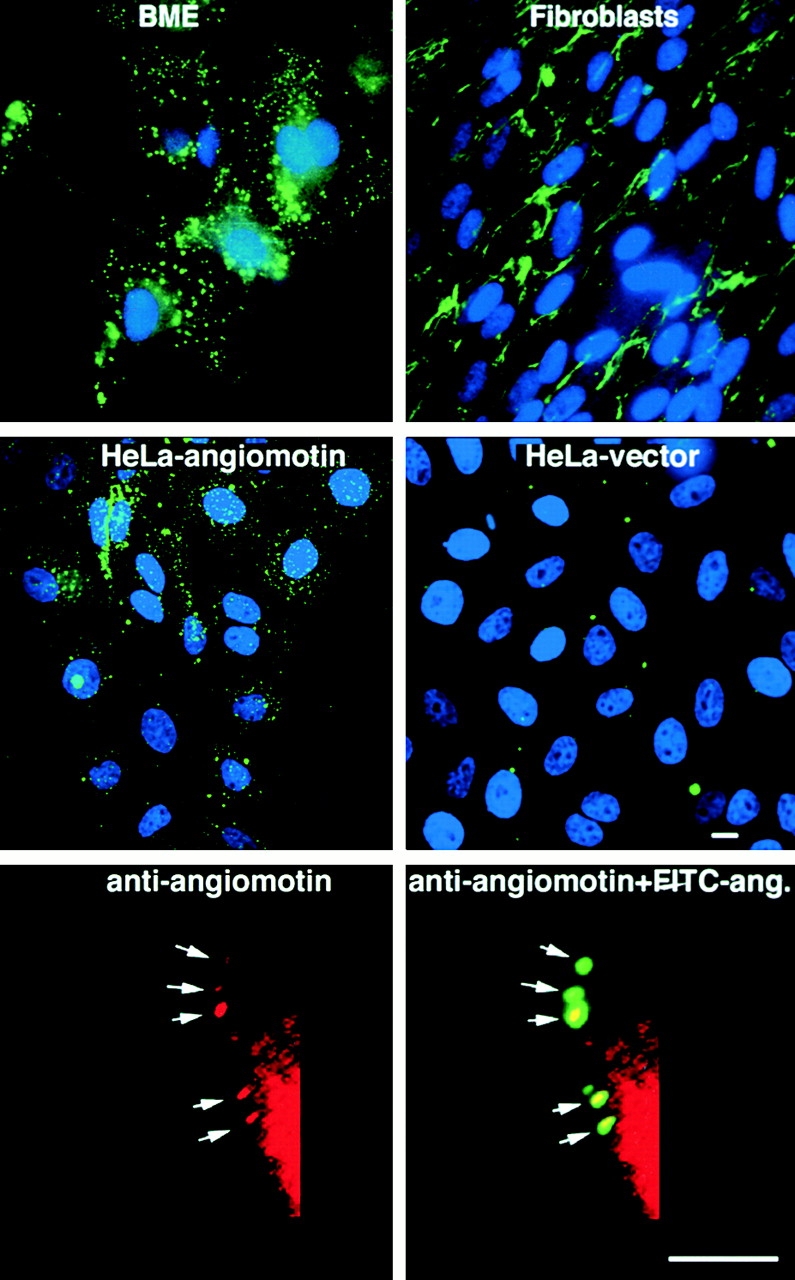
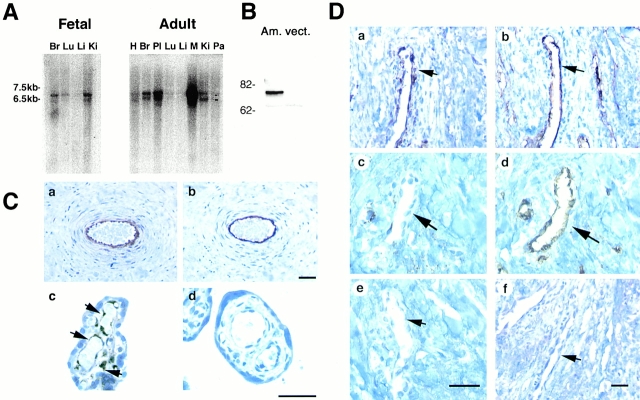
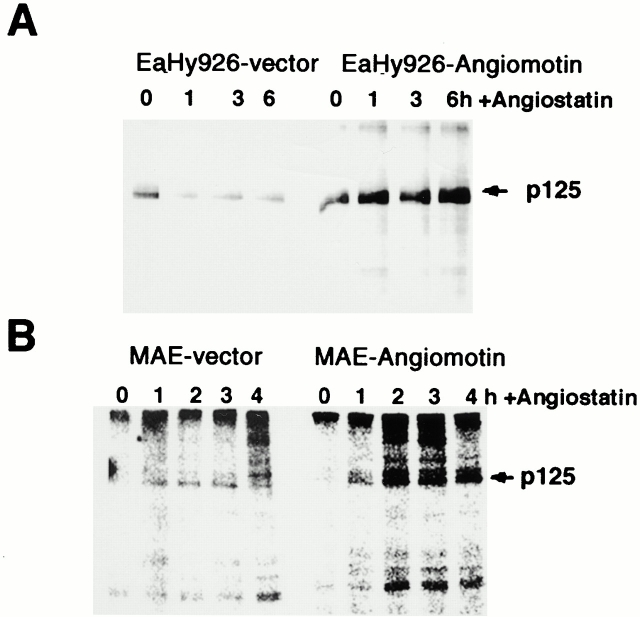
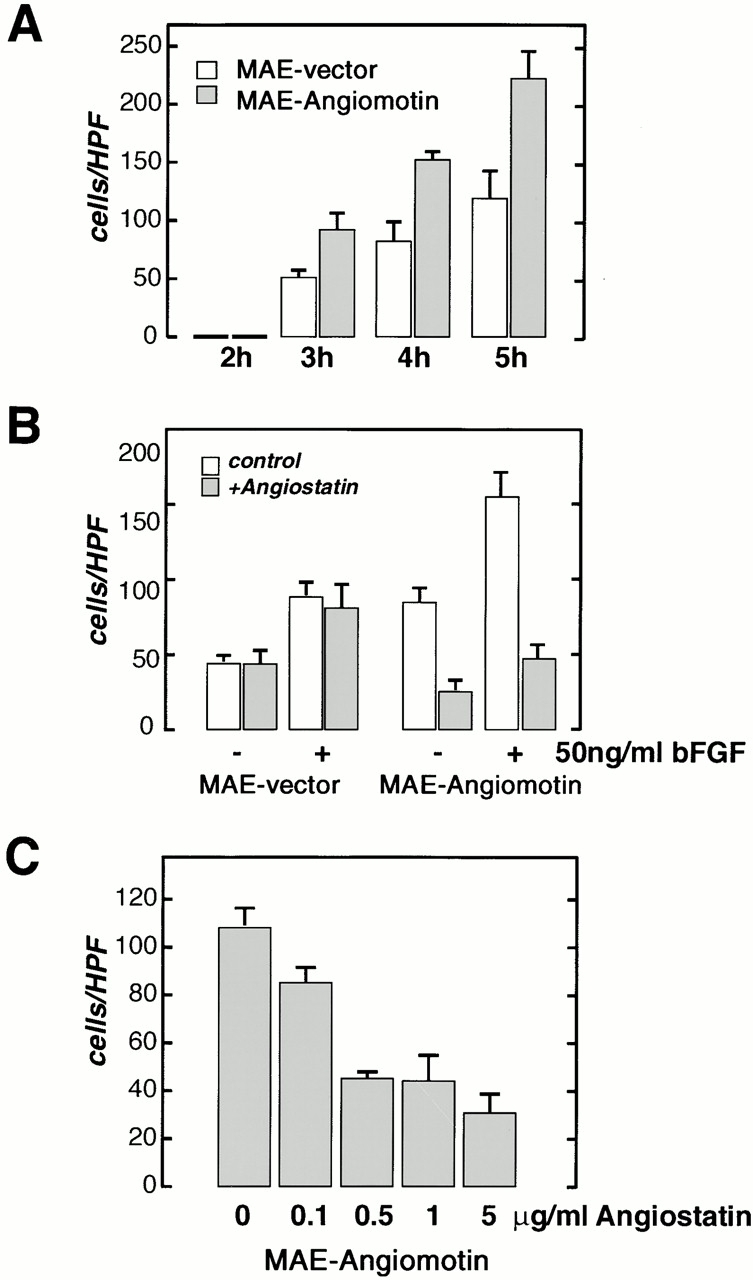
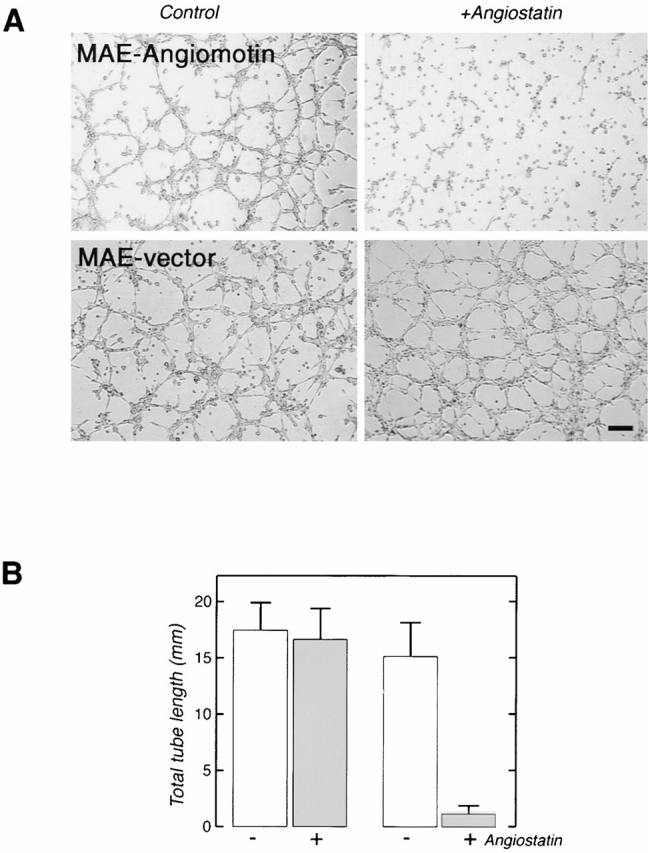
Comment in
-
Hold that line. Angiomotin regulates endothelial cell motility.J Cell Biol. 2001 Mar 19;152(6):F35-6. doi: 10.1083/jcb.152.6.f35. J Cell Biol. 2001. PMID: 11257132 Free PMC article. No abstract available.
References
-
- Ahmed A., Li X.F., Dunk C., Whittle M.J., Rushton D.I., Rollason T. Colocalisation of vascular endothelial growth factor and its Flt-1 receptor in human placenta. Growth Factors. 1995;12:235–243. - PubMed
-
- Barendsz-Janson A.F., Griffioen A.W., Muller A.D., van Dam-Mieras M.C., Hillen H.F. In vitro tumor angiogenesis assaysplasminogen lysine binding site 1 inhibits in vitro tumor-induced angiogenesis. J. Vasc. Res. 1998;35:109–114. - PubMed
-
- Bastaki M., Nelli E.E, Dell'Era P., Rusnati M., Molinari-Tosatti M.P., Parolini S., Auerbach R., Ruco L.P., Possati L., Presta M. Basic fibroblast growth factor-induced angiogenic phenotype in mouse endothelium. A study of aortic and microvascular endothelial cell lines. Arterioscler. Thromb. Vasc. Biol. 1997;17:454–464. - PubMed
-
- Bergers G., Javaherian K., Lo K.M., Folkman J., Hanahan D. Effects of angiogenesis inhibitors on multistage carcinogenesis in mice. Science. 1999;284:808–812. - PubMed
-
- Bouck N., Stellmach V., Hsu S.C. How tumors become angiogenic. Adv. Cancer Res. 1996;69:135–174. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
