

Evidence that gammadelta versus alphabeta T cell fate determination is initiated independently of T cell receptor signaling
- PMID: 11257136
- PMCID: PMC2193423
- DOI: 10.1084/jem.193.6.689
Evidence that gammadelta versus alphabeta T cell fate determination is initiated independently of T cell receptor signaling
Abstract
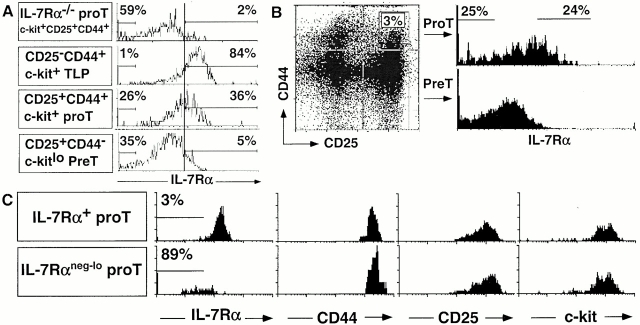
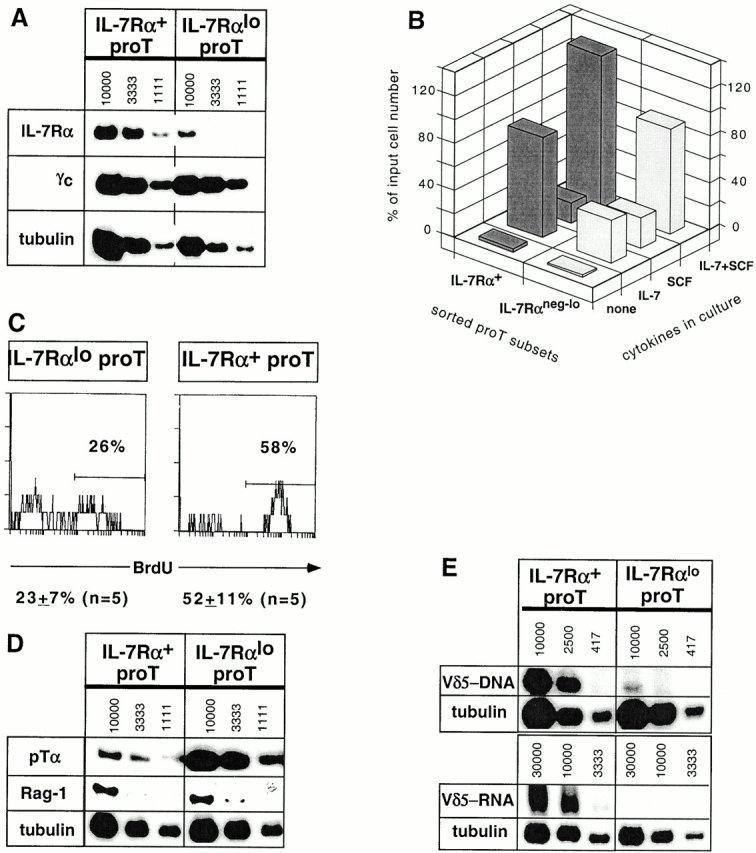
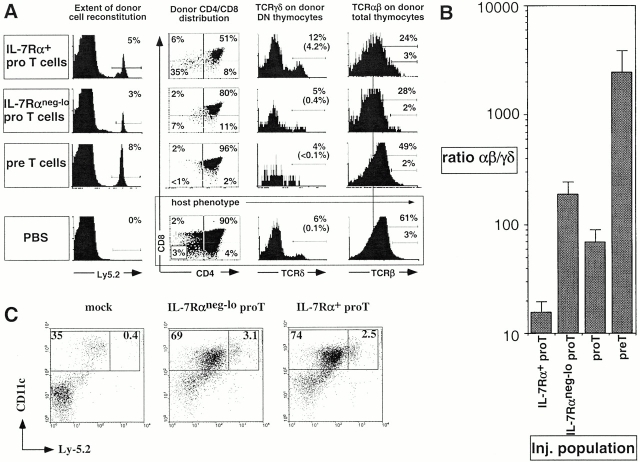
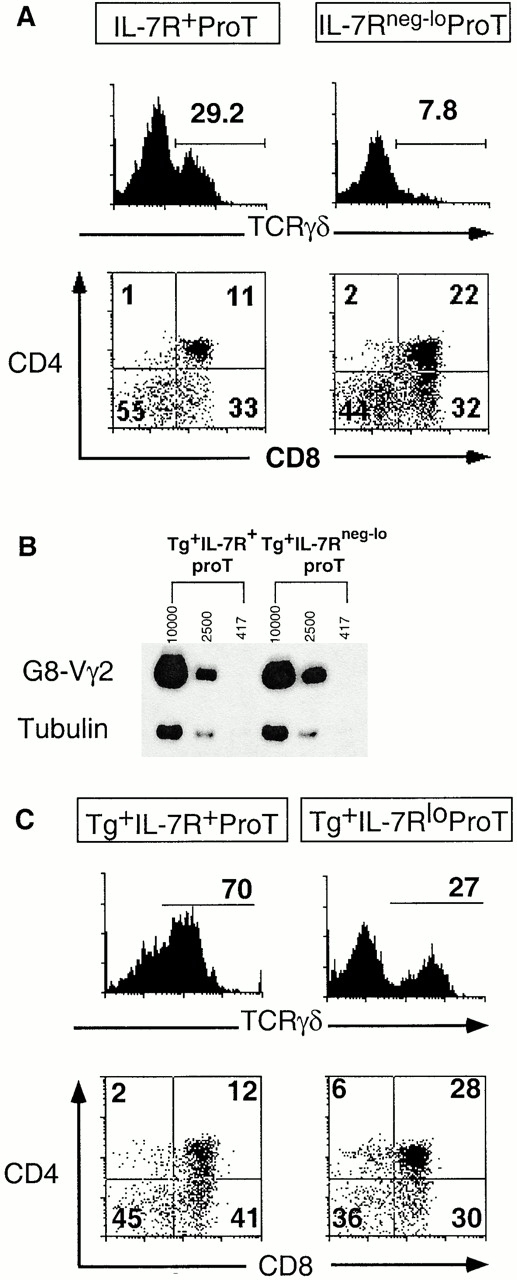
Two types of T cells, alphabeta and gammadelta, develop in vertebrates. How these two T cell lineages arise from a common thymic T progenitor is poorly understood. Differentiation of alphabeta lineage T cells requires the surrogate alpha chain (pTalpha), which associates with the T cell receptor (TCR) beta chain to form the pre-TCR. gammadelta lineage development does not appear to involve an obligatory surrogate chain, but instead requires productive rearrangement and expression of both TCR gamma and delta genes. It has been proposed that the quality of signals transmitted by the pre-TCR and gammadelta TCR are distinct and that these "instructive" signals determine the lineage fate of an uncommitted progenitor cell. Here we show that the thymic T progenitor cells (CD25(+)CD44(+)c-kit(+)CD3(-)CD4(-)CD8(-) thymocytes, termed pro-T cells) from young adult mice that have yet to express TCRs can be subdivided based on interleukin 7 receptor (IL-7R) expression. These subsets exhibit differential potential to develop into gammadelta versus alphabeta lineage (CD4+CD8+ cells) in the thymus. Upon intrathymic injection, IL-7R(neg-lo) pro-T cells generated a 13-fold higher ratio of alphabeta lineage to gammadelta lineage cells than did IL-7R(+) pro-T cells. Much of this difference was due to a fivefold greater potential of IL-7R(+) pro-T cells to develop into TCR-gammadelta T cells. Evidence indicates that this biased developmental potential is not a result of enhanced TCR-gamma gene rearrangement/expression in IL-7R(+) pro-T cells. These results indicate that the pro-T cells are heterogeneous in developmental potential before TCR gene rearrangement and suggest that in some precursor cells the initial lineage commitment is independent of TCR-mediated signals.
Figures
References
-
- Fehling H., Krotkova A., Saint-Ruf C., von Boehmer H. Crucial role of the pre-T cell receptor α gene in the development of αβ but not γδ T cells. Nature. 1995;375:795–798. - PubMed
-
- Dudley E.C., Petrie H.T., Shah L.M., Owen M.J., Hayday A.C. T cell receptor β chain gene rearrangement and selection during thymocyte development in adult mice. Immunity. 1994;1:83–93. - PubMed
-
- Penit C., Lucas B., Vasseur F. Cell expansion and growth arrest phases during the transition from precursor (CD4−8−) to immature (CD4+8+) thymocytes in normal and genetically modified mice. J. Immunol. 1995;154:5103–5113. - PubMed
-
- Kang J., Coles M., Cado D., Raulet D.H. The developmental fate of T cells is critically influenced by TCR γδ expression. Immunity. 1998;8:427–438. - PubMed
-
- Allison J.P., Lanier L.L. The T-cell antigen receptor γ generearrangement and cell lineages. Immunol. Today. 1987;8:293–296. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
