

Drosophila Mediator complex is broadly utilized by diverse gene-specific transcription factors at different types of core promoters
- PMID: 11259581
- PMCID: PMC86865
- DOI: 10.1128/MCB.21.7.2312-2323.2001
Drosophila Mediator complex is broadly utilized by diverse gene-specific transcription factors at different types of core promoters
Abstract
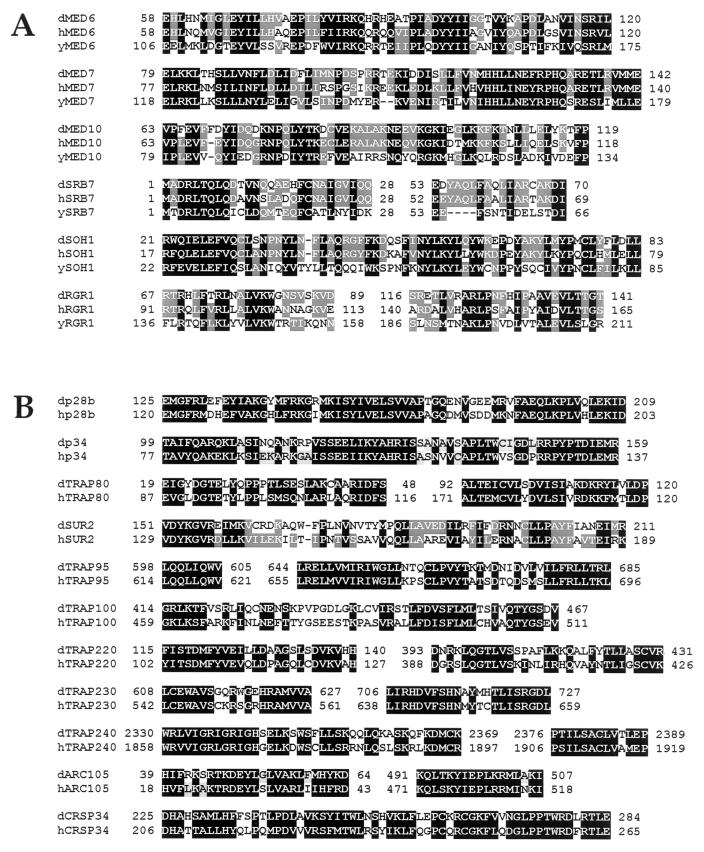
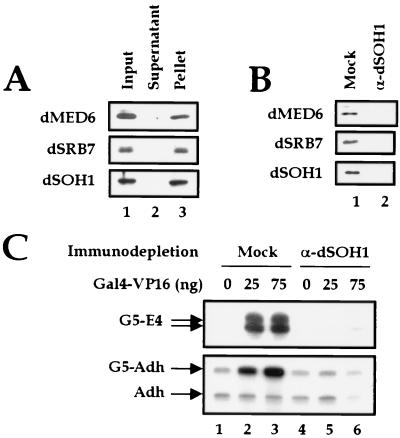
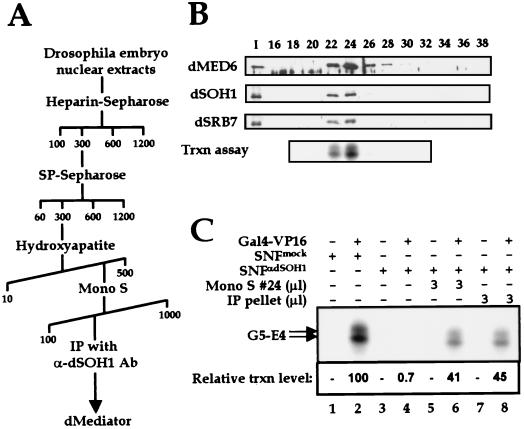
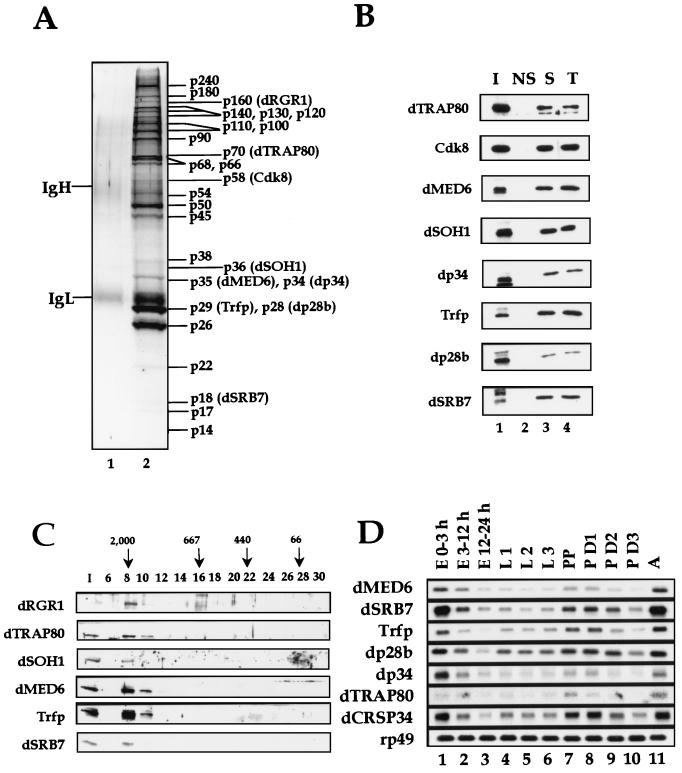
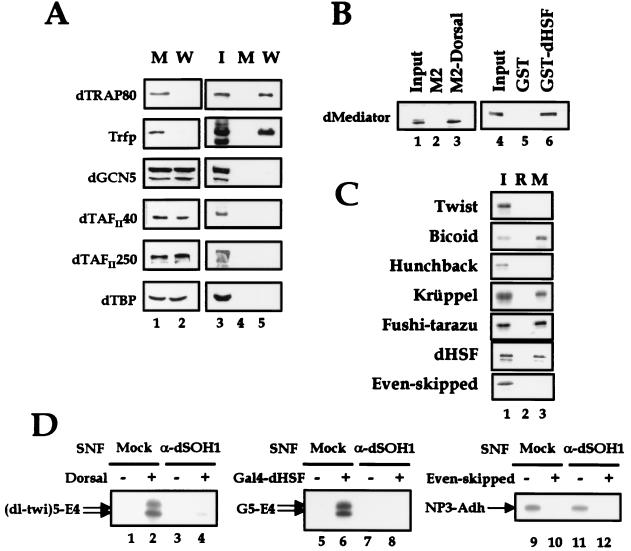
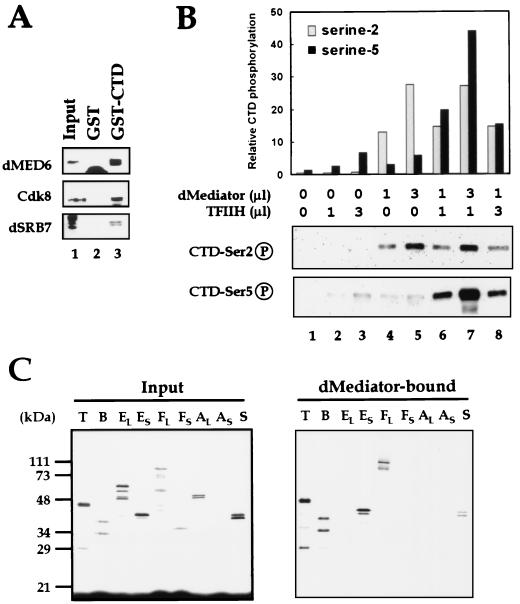
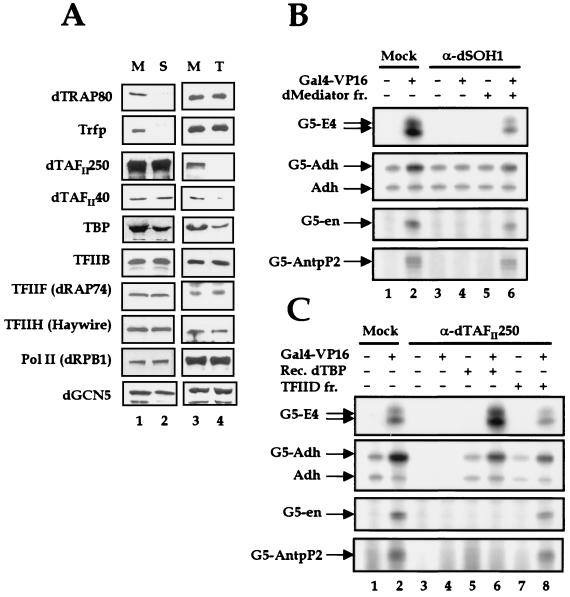
To decipher the mechanistic roles of Mediator proteins in regulating developmental specific gene expression and compare them to those of TATA-binding protein (TBP)-associated factors (TAFs), we isolated and analyzed a multiprotein complex containing Drosophila Mediator (dMediator) homologs. dMediator interacts with several sequence-specific transcription factors and basal transcription machinery and is critical for activated transcription in response to diverse transcriptional activators. The requirement for dMediator did not depend on a specific core promoter organization. By contrast, TAFs are preferentially utilized by promoters having a specific core element organization. Therefore, Mediator proteins are suggested to act as a pivotal coactivator that integrates promoter-specific activation signals to the basal transcription machinery.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases