

Mitochondrial translation of Saccharomyces cerevisiae COX2 mRNA is controlled by the nucleotide sequence specifying the pre-Cox2p leader peptide
- PMID: 11259585
- PMCID: PMC86869
- DOI: 10.1128/MCB.21.7.2359-2372.2001
Mitochondrial translation of Saccharomyces cerevisiae COX2 mRNA is controlled by the nucleotide sequence specifying the pre-Cox2p leader peptide
Abstract
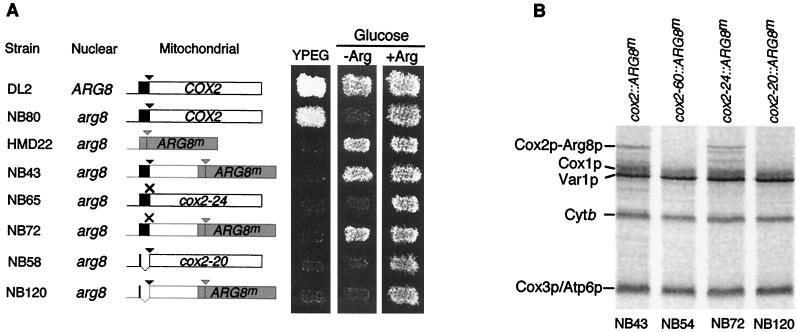
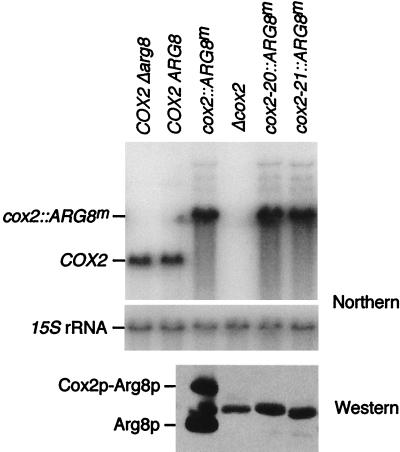
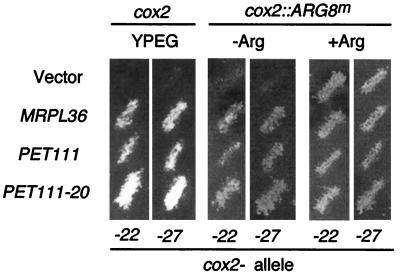
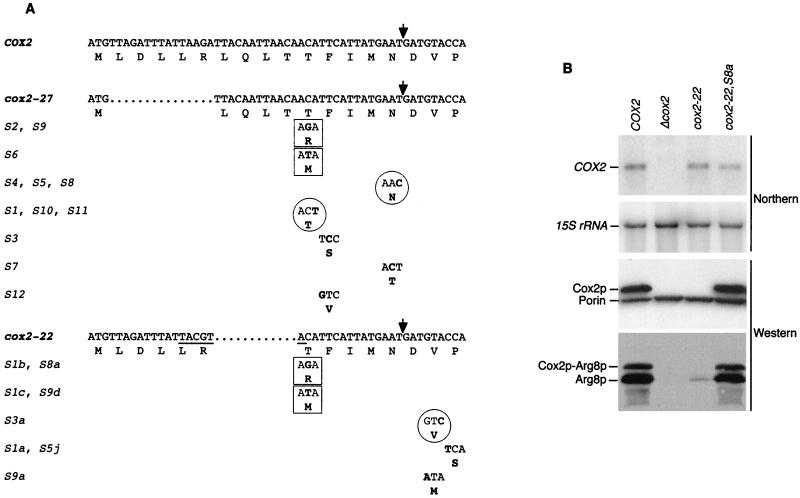
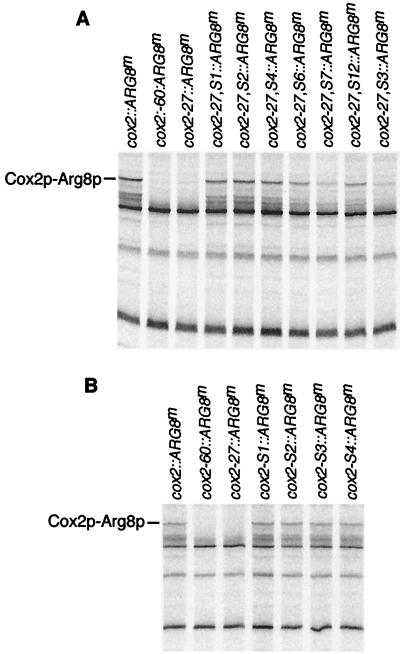
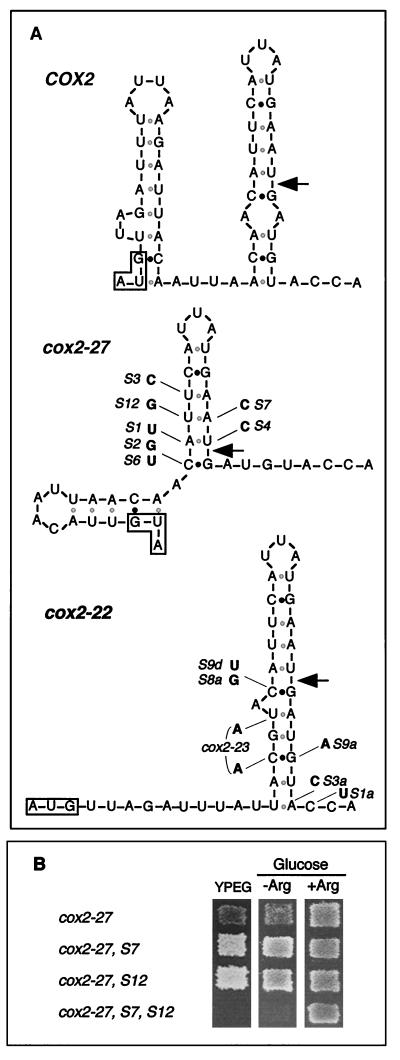
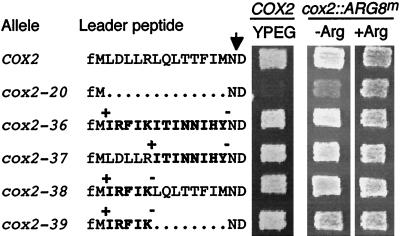
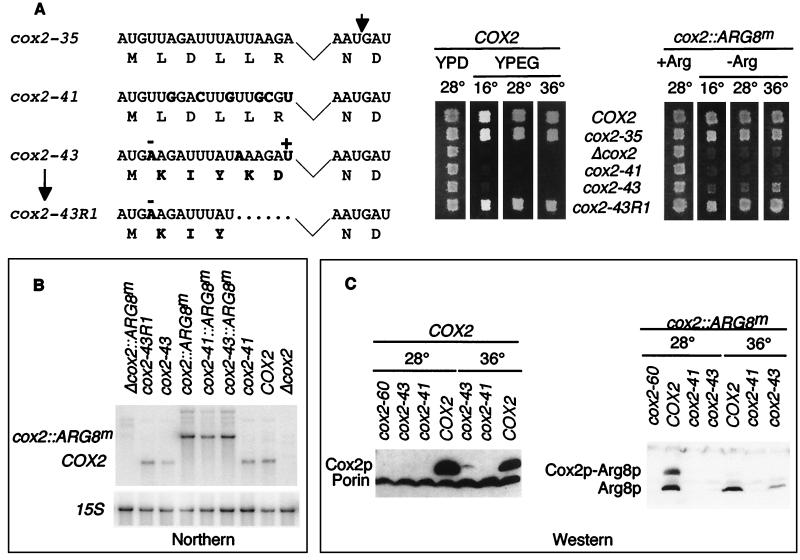
The mitochondrial gene encoding yeast cytochrome oxidase subunit II (Cox2p) specifies a precursor protein with a 15-amino-acid leader peptide. Deletion of the entire leader peptide coding region is known to block Cox2p accumulation posttranscriptionally. Here, we examined in vivo the role of the pre-Cox2p leader peptide and the mRNA sequence that encodes it in the expression of a mitochondrial reporter gene, ARG8m, fused to the 91st codon of COX2. We found within the coding sequence antagonistic elements that control translation: the positive element includes sequences in the first 14 codons specifying the leader peptide, while the negative element appears to be within codons 15 to 91. Partial deletions, point mutations, and local frameshifts within the leader peptide coding region were placed in both the cox2::ARG8m reporter and in COX2 itself. Surprisingly, the mRNA sequence of the first six codons specifying the leader peptide plays an important role in positively controlling translation, while the amino acid sequence of the leader peptide itself is relatively unconstrained. Two mutations that partially block translation can be suppressed by nearby sequence substitutions that weaken a predicted stem structure and by overproduction of either the COX2 mRNA-specific translational activator Pet111p or the large-subunit mitochondrial ribosomal protein MrpL36p. We propose that regulatory elements embedded in the translated COX2 mRNA sequence could play a role, together with trans-acting factors, in coupling regulated synthesis of nascent pre-Cox2p to its insertion in the mitochondrial inner membrane.
Figures

References
-
- Anderson D M, Schneewind O. A mRNA signal for the type III secretion of Yop proteins by Yersinia enterocolitica. Science. 1997;278:1140–1143. - PubMed
-
- Anderson S, De Bruijn M H L, Coulson A R, Eperon I C, Sanger F, Young I G. Complete sequence of bovine mitochondrial DNA: conserved features of the mammalian mitochondrial genome. J Mol Biol. 1982;156:683–717. - PubMed
-
- Behrens M, Michaelis G, Pratje E. Mitochondrial inner membrane protease 1 of Saccharomyces cerevisiae shows sequence similarity to the Escherichia coli leader peptidase. Mol Gen Genet. 1991;228:167–176. - PubMed
-
- Bonnefoy N, Chalvet F, Hamel P, Slonimski P P, Dujardin G. OXA1, a Saccharomyces cerevisiae nuclear gene whose sequence is conserved from prokaryotes to eukaryotes controls cytochrome oxidase biogenesis. J Mol Biol. 1994;239:201–212. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials