

Afferent input is necessary for seasonal growth and maintenance of adult avian song control circuits
- PMID: 11264307
- PMCID: PMC6762386
- DOI: 10.1523/JNEUROSCI.21-07-02320.2001
Afferent input is necessary for seasonal growth and maintenance of adult avian song control circuits
Abstract
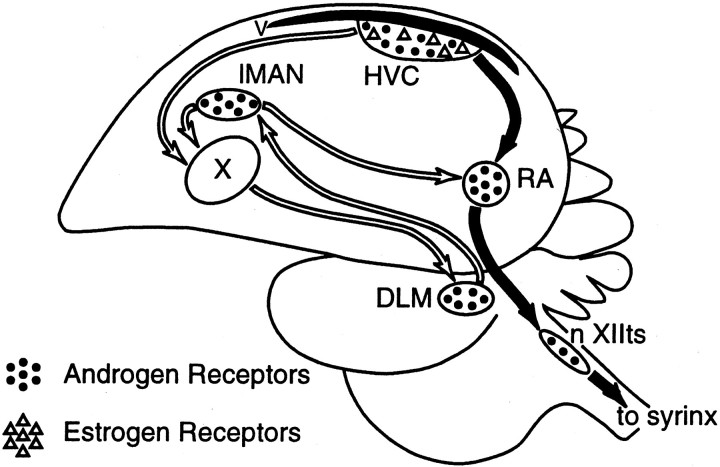
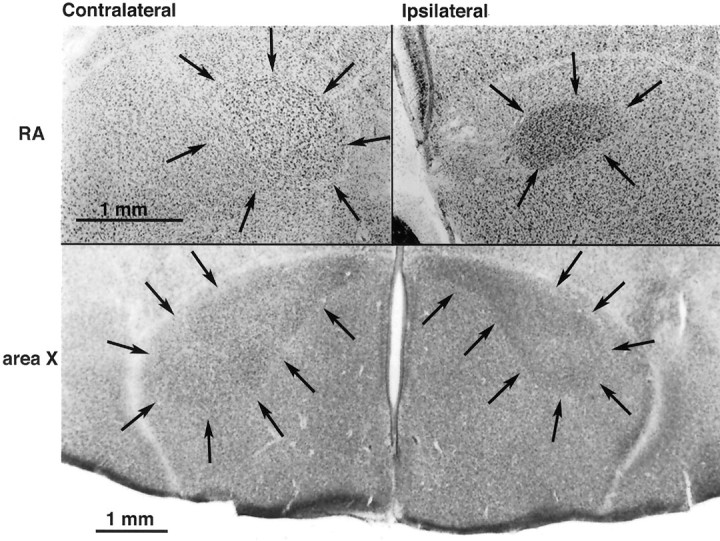
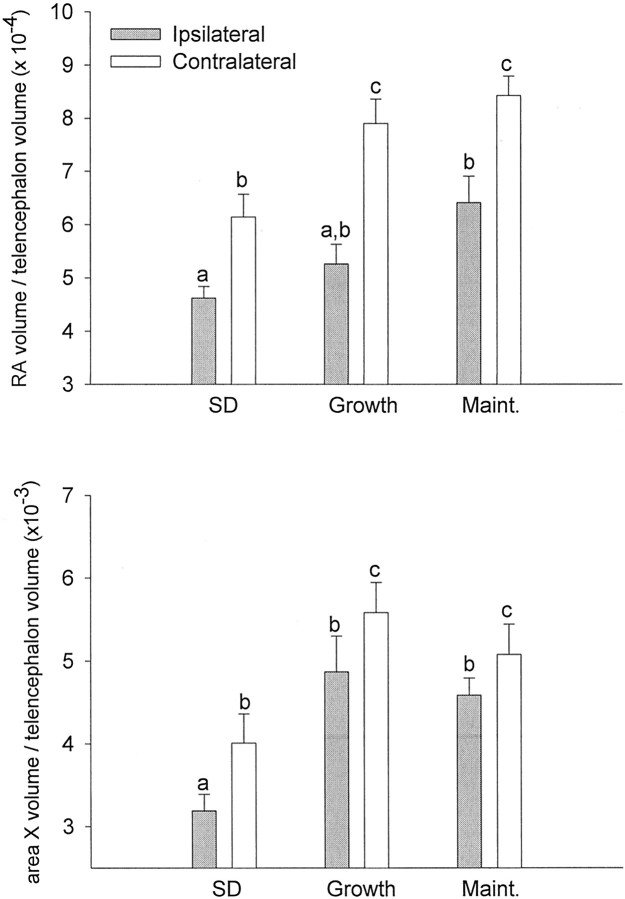
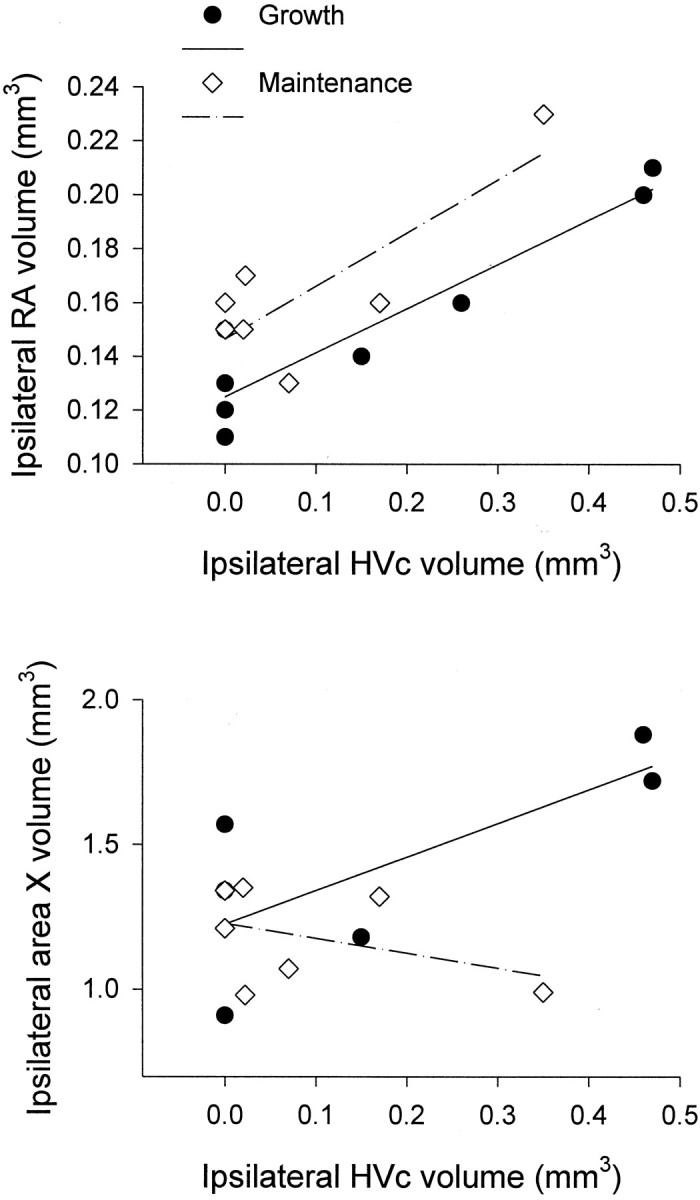
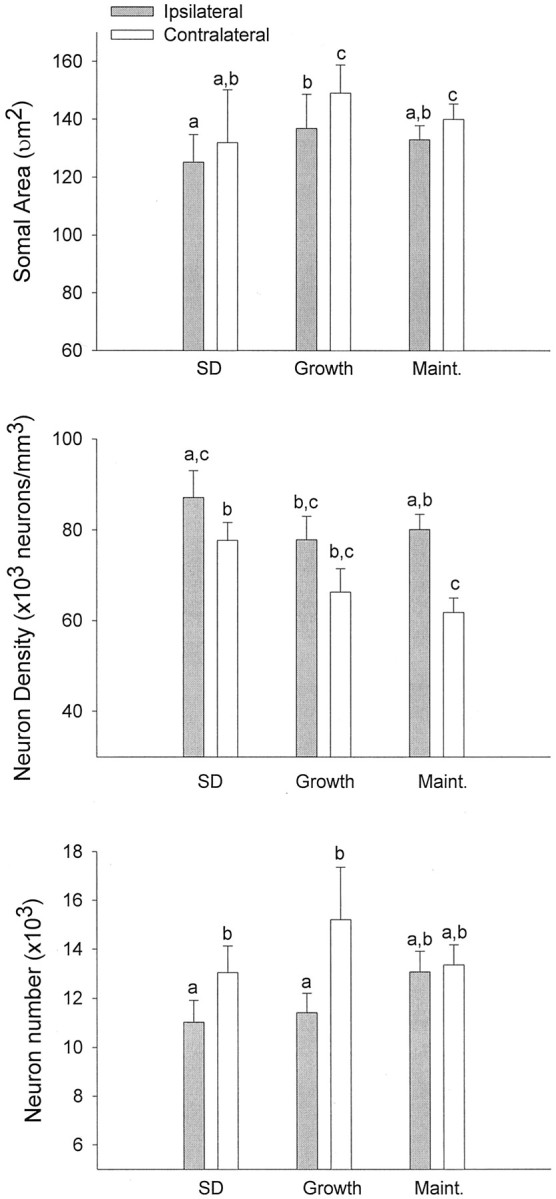
The neural circuits that regulate song behavior in adult songbirds undergo pronounced seasonal changes in morphology, primarily in response to changes in plasma testosterone (T). Most song nuclei have T receptors. We asked whether seasonal growth and maintenance of nuclei within these circuits are direct responses to the effects of T or its metabolites or are mediated indirectly via the effects of T on afferent nuclei. Photosensitive white-crowned sparrows were exposed to one of three treatments. (1) The neostriatal nucleus HVc (also known as the "high vocal center") was lesioned unilaterally, and the birds were exposed to long-day (LD) photoperiods and breeding levels of T for 30 d. (2) Birds were exposed to LD plus T (LD+T) for 30 d; then HVc was lesioned, and the birds were killed after an additional 30 d exposure to LD+T. (3) HVc was lesioned, and the sparrows were housed on short-day (SD) photoperiods in the absence of T treatment for 30 d. In both LD+T groups, the direct efferent targets of HVc, the robust nucleus of the archistriatum (RA) and area X, were smaller ipsilateral to the lesion. The lesion did not prevent growth of the hypoglossal motor nucleus, which does not receive direct afferent input from HVc. RA and area X were also smaller ipsilateral to the lesion in the SD birds. These results indicate that afferent input is required both for the growth of adult song circuits in response to typical breeding photoperiod and hormone conditions and for the maintenance of efferent nuclei in either their regressed or enlarged states.
Figures
References
-
- Arnold AP, Nottebohm F, Pfaff DW. Hormone concentrating cells in vocal control and other areas of the brain of the zebra finch (Poephila guttata). J Comp Neurol. 1976;165:487–511. - PubMed
-
- Balazs R, Hack N, Jorgensen OS, Cotman CW. N-methyl-d-aspartate promotes the survival of cerebellar granule cells: pharmacological characterization. Neurosci Lett. 1989;101:241–246. - PubMed
-
- Ball GF. Neuroendocrine basis of seasonal changes in vocal behavior among songbirds. In: Hauser M, Konishi M, editors. Neural mechanisms of communication. MIT; Cambridge, MA: 2000. pp. 213–253.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials