

Antiviral effect of N-butyldeoxynojirimycin against bovine viral diarrhea virus correlates with misfolding of E2 envelope proteins and impairment of their association into E1-E2 heterodimers
- PMID: 11264342
- PMCID: PMC114844
- DOI: 10.1128/JVI.75.8.3527-3536.2001
Antiviral effect of N-butyldeoxynojirimycin against bovine viral diarrhea virus correlates with misfolding of E2 envelope proteins and impairment of their association into E1-E2 heterodimers
Abstract
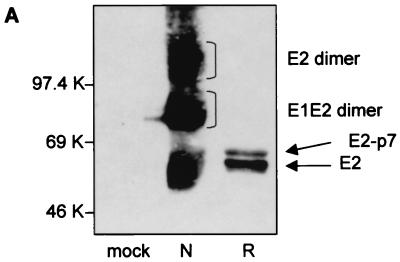
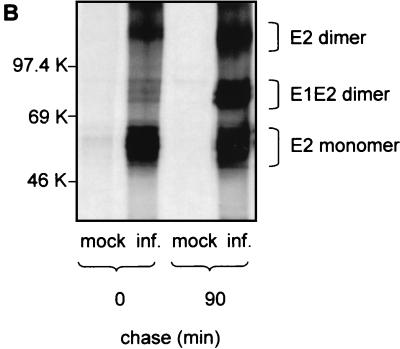
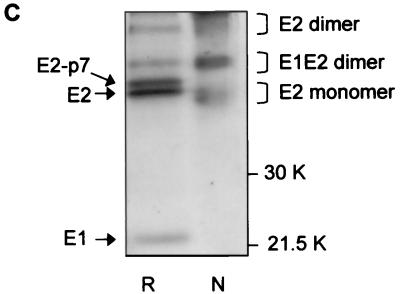
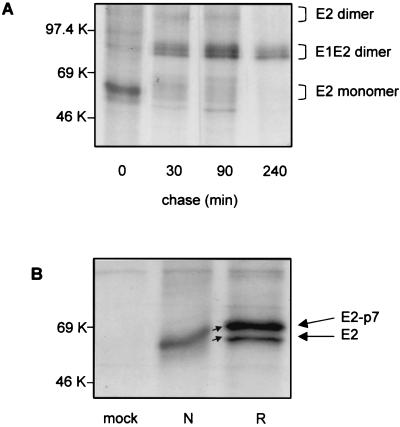
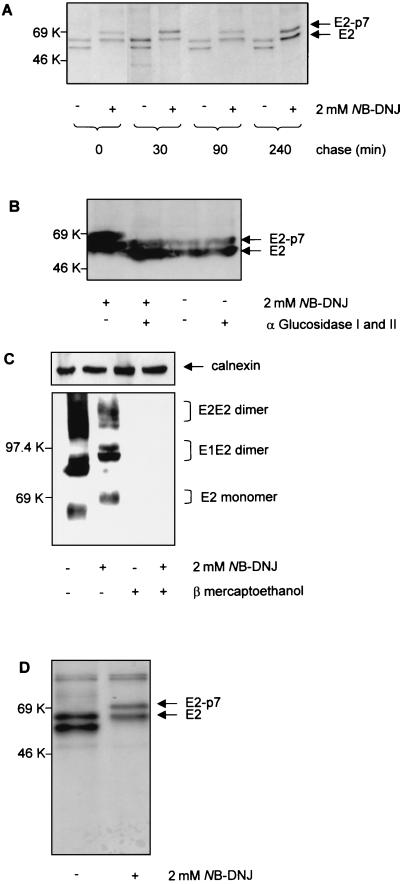
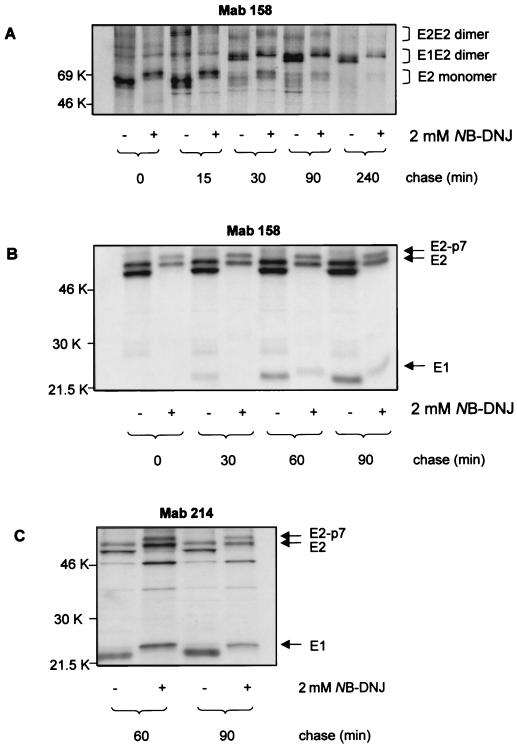
The iminosugar N-butyldeoxynojirimycin (NB-DNJ), an endoplasmic reticulum alpha-glucosidase inhibitor, has an antiviral effect against bovine viral diarrhea virus (BVDV). In this report, we investigate the molecular mechanism of this inhibition by studying the folding pathway of BVDV envelope glycoproteins in the presence and absence of NB-DNJ. Our results show that, while the disulfide-dependent folding of E2 glycoprotein occurs rapidly (2.5 min), the folding of E1 occurs slowly (30 min). Both BVDV envelope glycoproteins associate rapidly with calnexin and dissociate with different kinetics. The release of E1 from the interaction with calnexin coincides with the beginning of E1 and E2 association into disulfide-linked heterodimers. In the presence of NB-DNJ, the interaction of E1 and E2 with calnexin is prevented, leading to misfolding of the envelope glycoproteins and inefficient formation of E1-E2 heterodimers. The degree of misfolding and the lack of association of E1 and E2 into disulfide-linked complexes in the presence of NB-DNJ correlate with the dose-dependent antiviral effect observed for this iminosugar.
Figures


References
-
- Bergeron J J M, Brenner M B, Thomas D Y, Williams D B. Calnexin: a membrane-bound chaperone of the endoplasmic reticulum. Trends Biochem Sci. 1994;19:124–129. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
