

Characterization of uracil-DNA glycosylase activity from Trypanosoma cruzi and its stimulation by AP endonuclease
- PMID: 11266557
- PMCID: PMC31275
- DOI: 10.1093/nar/29.7.1549
Characterization of uracil-DNA glycosylase activity from Trypanosoma cruzi and its stimulation by AP endonuclease
Abstract
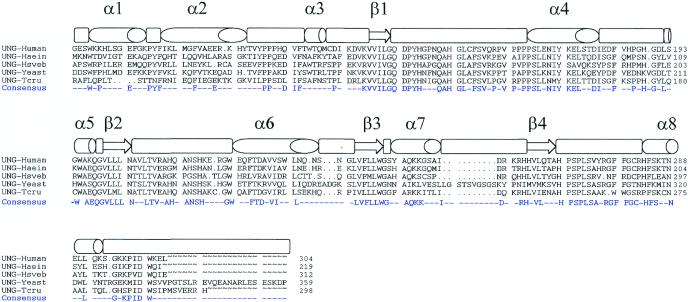
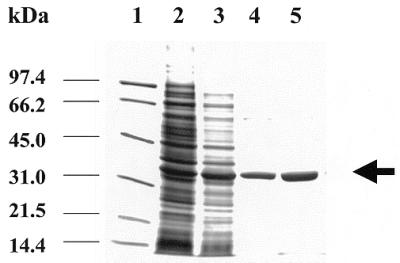
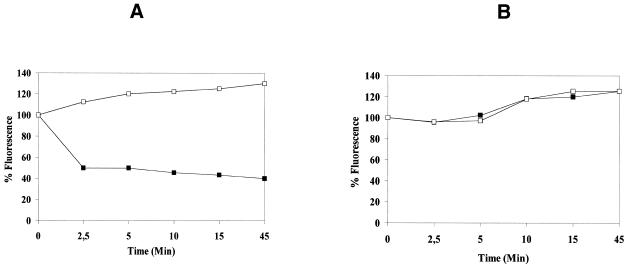
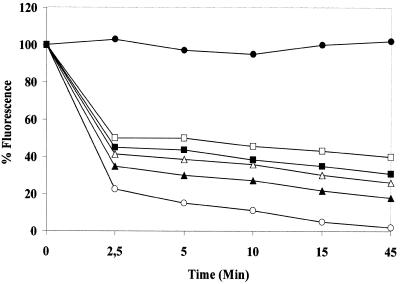
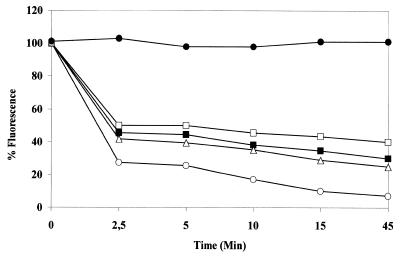
The intracellular pathogen Trypanosoma cruzi is the etiological agent of Chagas' disease. We have isolated a full-length cDNA encoding uracil-DNA glycosylase (UDGase), a key enzyme involved in DNA repair, from this organism. The deduced protein sequence is highly conserved at the C-terminus of the molecule and shares key residues involved in binding or catalysis with most of the UDGases described so far, while the N-terminal part is highly variable. The gene is single copy and is located on a chromosome of approximately 1.9 Mb. A His-tagged recombinant protein was overexpressed, purified and used to raise polyclonal antibodies. Western blot analysis revealed the existence of a single UDGase species in parasite extracts. Using a specific ethidium bromide fluorescence assay, recombinant T.cruzi UDGase was shown to specifically excise uracil from DNA. The addition of both Leishmania major AP endonuclease and exonuclease III, the major AP endonuclease from Escherichia coli, produces stimulation of UDGase activity. This activation is specific for AP endonuclease and suggests functional communication between the two enzymes.
Figures

Similar articles
-
Apurinic/apyrimidinic endonuclease genes from the trypanosomatidae leishmania major and Trypanosoma cruzi confer resistance to oxidizing agents in DNA repair-deficient Escherichia coli.Nucleic Acids Res. 1999 Feb 1;27(3):771-7. doi: 10.1093/nar/27.3.771. Nucleic Acids Res. 1999. PMID: 9889272 Free PMC article.
-
Enhanced activity of adenine-DNA glycosylase (Myh) by apurinic/apyrimidinic endonuclease (Ape1) in mammalian base excision repair of an A/GO mismatch.Nucleic Acids Res. 2001 Feb 1;29(3):743-52. doi: 10.1093/nar/29.3.743. Nucleic Acids Res. 2001. PMID: 11160897 Free PMC article.
-
cDNA cloning, expression and functional characterization of an Arabidopsis thaliana homologue of the Escherichia coli DNA repair enzyme endonuclease III.Plant Mol Biol. 2000 Sep;44(1):43-52. doi: 10.1023/a:1006429114451. Plant Mol Biol. 2000. PMID: 11094978
-
Lessons learned from structural results on uracil-DNA glycosylase.Mutat Res. 2000 Aug 30;460(3-4):183-99. doi: 10.1016/s0921-8777(00)00026-4. Mutat Res. 2000. PMID: 10946228 Review.
-
Molecular mechanism of PCNA-dependent base excision repair.Prog Nucleic Acid Res Mol Biol. 2001;68:129-38. doi: 10.1016/s0079-6603(01)68095-4. Prog Nucleic Acid Res Mol Biol. 2001. PMID: 11554292 Review.
Cited by
-
Expression and the Peculiar Enzymatic Behavior of the Trypanosoma cruzi NTH1 DNA Glycosylase.PLoS One. 2016 Jun 10;11(6):e0157270. doi: 10.1371/journal.pone.0157270. eCollection 2016. PLoS One. 2016. PMID: 27284968 Free PMC article.
-
Ma-LMM01 infecting toxic Microcystis aeruginosa illuminates diverse cyanophage genome strategies.J Bacteriol. 2008 Mar;190(5):1762-72. doi: 10.1128/JB.01534-07. Epub 2007 Dec 7. J Bacteriol. 2008. PMID: 18065537 Free PMC article.
-
Differential modes of DNA binding by mismatch uracil DNA glycosylase from Escherichia coli: implications for abasic lesion processing and enzyme communication in the base excision repair pathway.Nucleic Acids Res. 2011 Apr;39(7):2593-603. doi: 10.1093/nar/gkq913. Epub 2010 Nov 25. Nucleic Acids Res. 2011. PMID: 21112870 Free PMC article.
-
DNA repair pathways in trypanosomatids: from DNA repair to drug resistance.Microbiol Mol Biol Rev. 2014 Mar;78(1):40-73. doi: 10.1128/MMBR.00045-13. Microbiol Mol Biol Rev. 2014. PMID: 24600040 Free PMC article. Review.
-
Overview of DNA Repair in Trypanosoma cruzi, Trypanosoma brucei, and Leishmania major.J Nucleic Acids. 2010 Oct 4;2010:840768. doi: 10.4061/2010/840768. J Nucleic Acids. 2010. PMID: 20976268 Free PMC article.
References
-
- Nicholl I.D., Nealon,K. and Kenny,M.K. (1997) Reconstitution of human base excision repair with purified proteins. Biochemistry, 36, 7557–7566. - PubMed
-
- Mauro D.J., De Reil,J.K., Tallarida,R.J. and Sirover,M.A. (1993) Mechanisms of excision of 5-fluoracil by uracil DNA glycosylase in normal human cells. Mol. Pharmacol., 43, 854–857. - PubMed
-
- Hatahet Z., Kow,Y.W., Purmal,A.A., Cunningham,R.P. and Wallace,S.S. (1994) New substrates for old enzymes. 5-Hydroxy-2′-deoxycytidine and 5-hydroxy-2′-deoxyuridine are substrates for Escherichia coli endonuclease III and formamidopyrimidine DNA. J. Biol. Chem., 269, 18814–18820. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Miscellaneous
