

Ulex europaeus agglutinin II (UEA-II) is a novel, potent inhibitor of complement activation
- PMID: 11266613
- PMCID: PMC2373946
- DOI: 10.1110/ps.26401
Ulex europaeus agglutinin II (UEA-II) is a novel, potent inhibitor of complement activation
Abstract
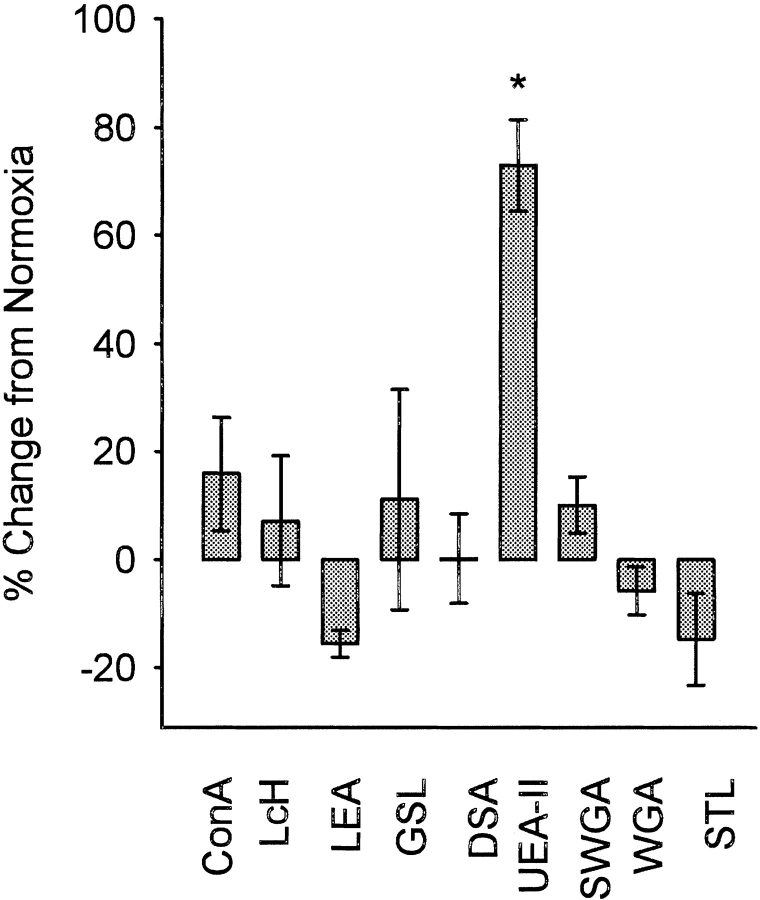
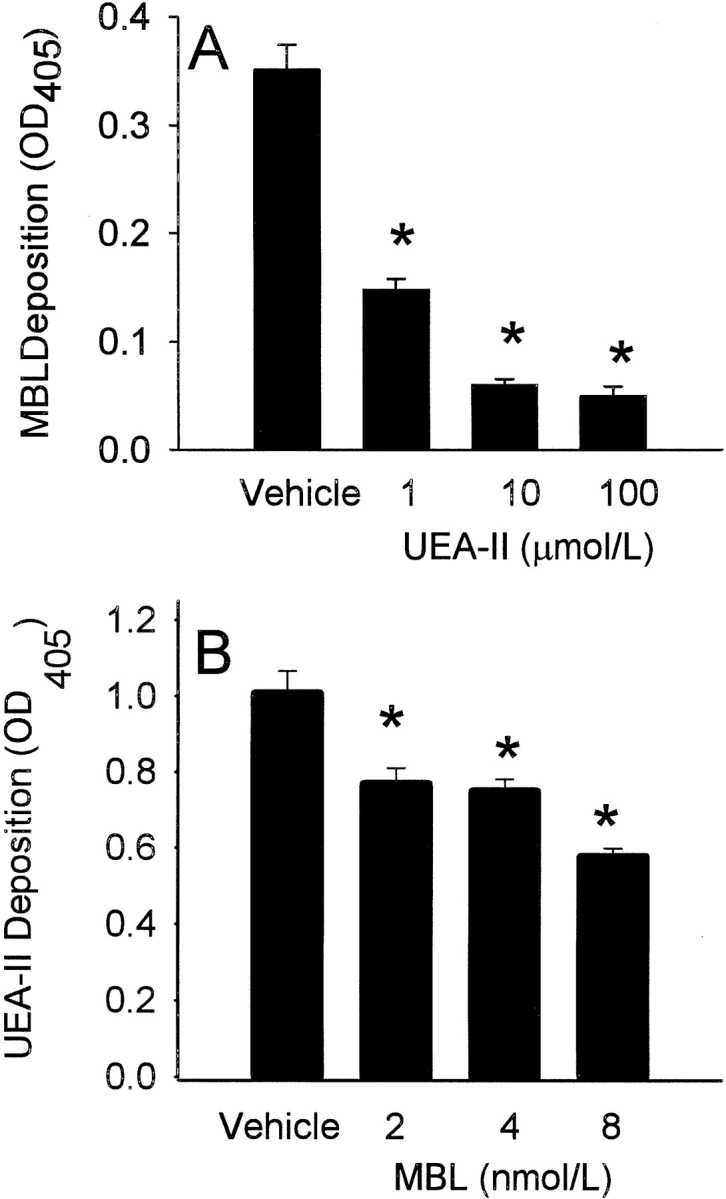
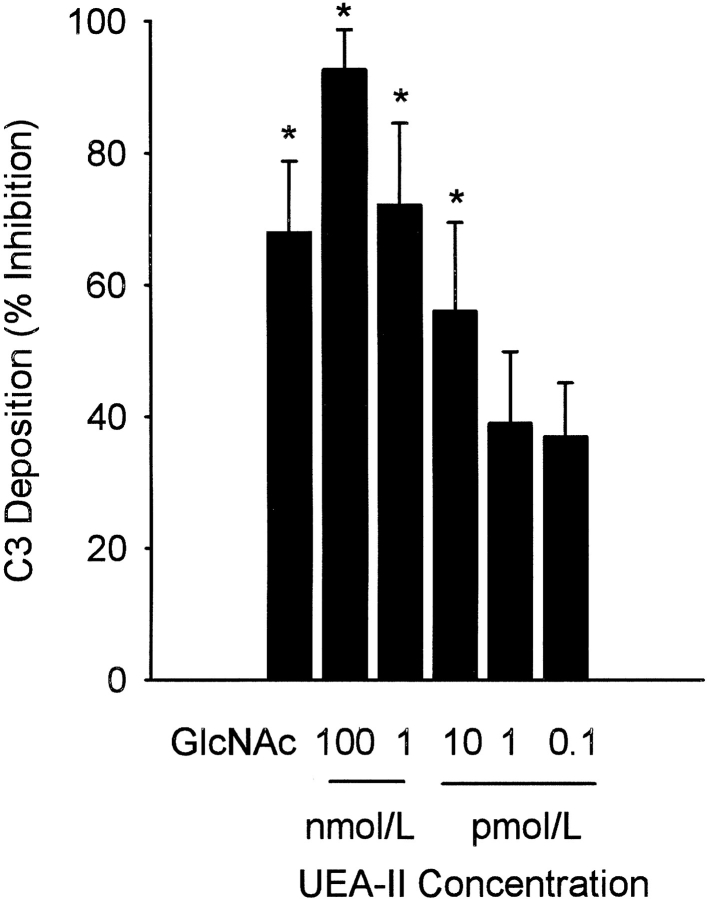
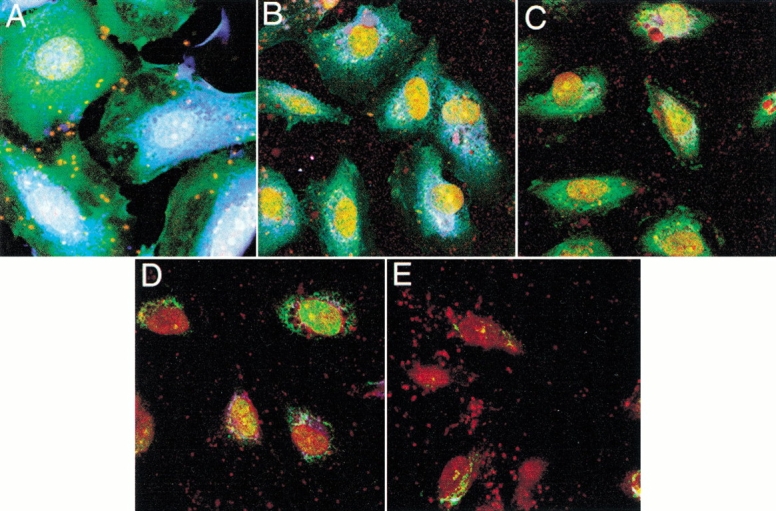
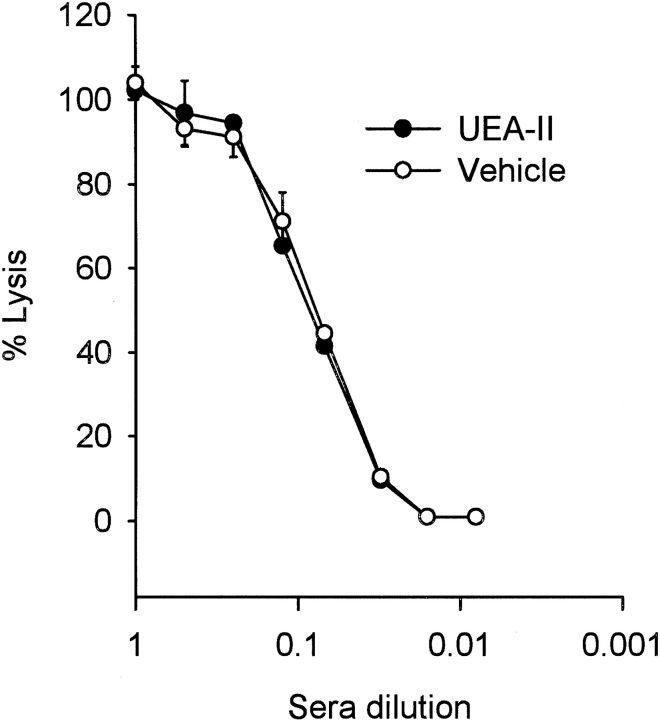
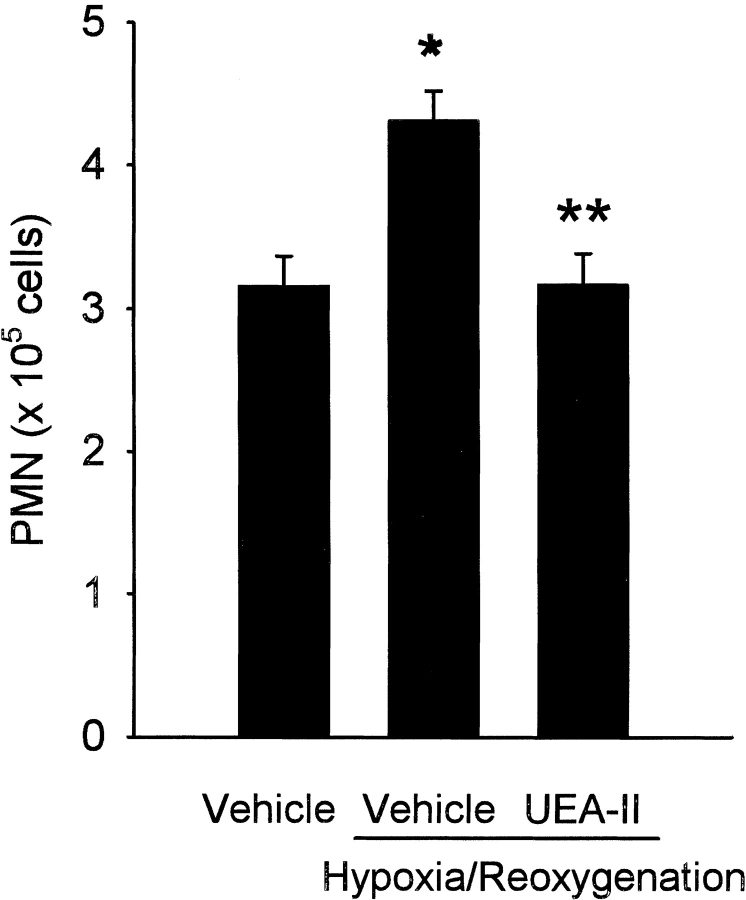
Complement is an important mediator of vascular injury following oxidative stress. We recently demonstrated that complement activation following endothelial oxidative stress is mediated by mannose-binding lectin (MBL) and activation of the lectin complement pathway. Here, we investigated whether nine plant lectins which have a binding profile similar to that of MBL competitively inhibit MBL deposition and subsequent complement activation following human umbilical vein endothelial cell (HUVEC) oxidative stress. HUVEC oxidative stress (1% O(2), 24 hr) significantly increased Ulex europaeus agglutinin II (UEA-II) binding by 72 +/- 9% compared to normoxic cells. UEA-II inhibited MBL binding to HUVEC in a concentration-dependent manner following oxidative stress. Further, MBL inhibited UEA-II binding to HUVEC in a concentration-dependent manner following oxidative stress, suggesting a common ligand. UEA-II (< or = 100 micromol/L) did not attenuate the hemolytic activity, nor did it inhibit C3a des Arg formation from alternative or classical complement pathway-specific hemolytic assays. C3 deposition (measured by ELISA) following HUVEC oxidative stress was inhibited by UEA-II in a concentration-dependent manner (IC(50) = 10 pmol/L). UEA-II inhibited C3 and MBL co-localization (confocal microscopy) in a concentration-dependent manner on HUVEC following oxidative stress (IC(50) approximately 1 pmol/L). Finally, UEA-II significantly inhibited complement-dependent neutrophil chemotaxis, but failed to inhibit fMLP-mediated chemotaxis, following endothelial oxidative stress. These data demonstrate that UEA-II is a novel, potent inhibitor of human MBL deposition and complement activation following human endothelial oxidative stress.
Figures
Similar articles
-
Endothelial oxidative stress activates the lectin complement pathway: role of cytokeratin 1.Am J Pathol. 2001 Sep;159(3):1045-54. doi: 10.1016/S0002-9440(10)61779-8. Am J Pathol. 2001. PMID: 11549596 Free PMC article.
-
A keratin peptide inhibits mannose-binding lectin.J Immunol. 2001 Mar 15;166(6):4148-53. doi: 10.4049/jimmunol.166.6.4148. J Immunol. 2001. PMID: 11238665
-
A novel measurement method for activation of the lectin complement pathway via both mannose-binding lectin (MBL) and L-ficolin.J Immunol Methods. 2009 Sep 30;349(1-2):9-17. doi: 10.1016/j.jim.2009.08.005. Epub 2009 Aug 20. J Immunol Methods. 2009. PMID: 19699205
-
New perspectives on mannan-binding lectin-mediated complement activation.Immunobiology. 2007;212(4-5):301-11. doi: 10.1016/j.imbio.2006.12.004. Epub 2007 Jan 22. Immunobiology. 2007. PMID: 17544815 Review.
-
Complement activation following oxidative stress.Mol Immunol. 1999 Sep-Oct;36(13-14):941-8. doi: 10.1016/s0161-5890(99)00116-9. Mol Immunol. 1999. PMID: 10698348 Review.
Cited by
-
Endothelial oxidative stress activates the lectin complement pathway: role of cytokeratin 1.Am J Pathol. 2001 Sep;159(3):1045-54. doi: 10.1016/S0002-9440(10)61779-8. Am J Pathol. 2001. PMID: 11549596 Free PMC article.
-
Complement activation and inhibition in wound healing.Clin Dev Immunol. 2012;2012:534291. doi: 10.1155/2012/534291. Epub 2012 Dec 30. Clin Dev Immunol. 2012. PMID: 23346185 Free PMC article. Review.
-
The complement system in ischemia-reperfusion injuries.Immunobiology. 2012 Nov;217(11):1026-33. doi: 10.1016/j.imbio.2012.07.024. Epub 2012 Aug 7. Immunobiology. 2012. PMID: 22964228 Free PMC article. Review.
-
A Useful Guide to Lectin Binding: Machine-Learning Directed Annotation of 57 Unique Lectin Specificities.ACS Chem Biol. 2022 Nov 18;17(11):2993-3012. doi: 10.1021/acschembio.1c00689. Epub 2022 Jan 27. ACS Chem Biol. 2022. PMID: 35084820 Free PMC article.
References
-
- Bless, N.M., Warner, R.L., Padgaonkar, V.A., Lentsch, A.B., Czermak, B.J., Schmal, H., Friedl, H.P., and Ward, P.A. 1999. Roles for C-X-C chemokines and C5a in lung injury after hindlimb ischemia-reperfusion. Am J Physiol 276 L57–L63. - PubMed
-
- Buerke, M., Prüfer, D., Dahm, M., Oelert, H., Meyer, J., and Darius, H. 1998. Blocking of classical complement pathway inhibits endothelial adhesion molecule expression and preserves ischemic myocardium from reperfusion injury. Journal of Pharmacology and Experimental Therapeutics 286 429–438. - PubMed
-
- Collard, C.D., Agah, A., Reenstra, W.R., Buras, J., and Stahl, G.L. 1999. Endothelial nuclear factor-kB translocation and vascular cell adhesion molecule-1 induction by complement: Inhibition with anti-human C5 therapy or cGMP analogues. Arterioscler Thromb Vasc Biol 19 2623–2629. - PubMed
-
- Collard, C.D., Vakeva, A., Bukusoglu, C., Zund, G., Sperati, C.J., Colgan, S.P., and Stahl, G.L. 1997. Reoxygenation of hypoxic human umbilical vein endothelial cells (HUVECs) activates the classic complement pathway. Circulation 96 326–333. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
