

doi: 10.1128/JB.183.8.2439-2444.2001.
Independence of circadian timing from cell division in cyanobacteria
Affiliations
- PMID: 11274102
- PMCID: PMC95159
- DOI: 10.1128/JB.183.8.2439-2444.2001
Item in Clipboard
Independence of circadian timing from cell division in cyanobacteria
J Bacteriol.
2001 Apr.
Abstract
In the cyanobacterium Synechococcus elongatus, cell division is regulated by a circadian clock. Deletion of the circadian clock gene, kaiC, abolishes rhythms of gene expression and cell division timing. Overexpression of the ftsZ gene halted cell division but not growth, causing cells to grow as filaments without dividing. The nondividing filamentous cells still exhibited robust circadian rhythms of gene expression. This result indicates that the circadian timing system is independent of rhythmic cell division and, together with other results, suggests that the cyanobacterial circadian system is stable and well sustained under a wide range of intracellular conditions.
Figures
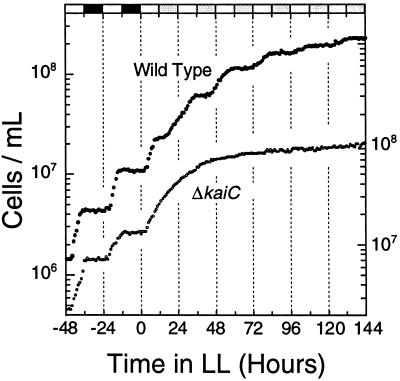
Circadian rhythm of cell division in batch cultures of S. elongatus. Cell number data for the wild-type strain (●) and a clock-null ΔkaiC strain (○). The wild-type and ΔkaiC (46) strains were grown in LD 12:12 and transferred into LL (45 μE/m2/s) at time zero. The last two LD cycles preceding LL are illustrated by the bars on the upper abscissa (white, light; black, darkness; gray, subjective night phases of LL). The left ordinate is for the wild-type strain, and the right ordinate is for the ΔkaiC strain.
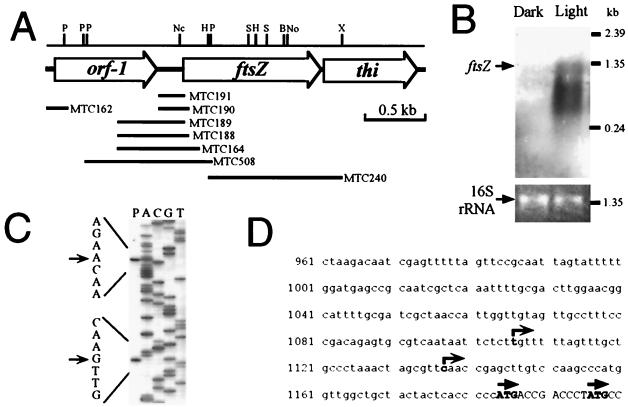
Primary structure and expression of the S. elongatus ftsZ gene. (A) Physical map of the ftsZ gene and its flanking regions. Segments used for constructing luxAB reporter strains are indicated by horizontal bars with the names of the transformed strains. P, PstI; Nc, NcoI; H, HindIII; S, SalI; B, BssHI; No, NotI; X, XhoI. (B) Northern blot analysis of the ftsZ gene. To show equivalent loading of the two lanes in the gel, ethidium bromide staining of the 16S rRNA band is shown. Cells were grown in LD 12:12 cycles and harvested in the day and night. In LD cycles, cyanobacteria divide only in the day (Fig. 1). (C) Determination of putative transcriptional start sites of the ftsZ gene by primer extension. The transcription start site was confirmed by S1 mapping (not shown). (D) Sequence of the promoter region of ftsZ. Putative transcriptional (right-angle arrows) and translational (straight arrows) start sites are indicated.
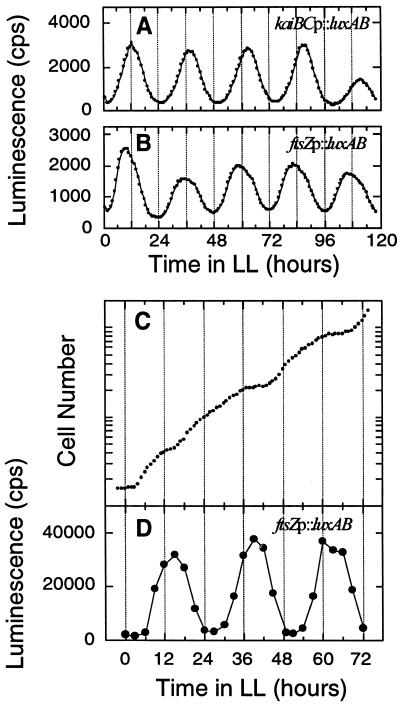
Rhythms of luminescence in ftsZp::luxAB reporter strain MTC508. (A and B) In vivo luminescence from 3-ml batch liquid cultures of kaiBCp::luxAB (A) and ftsZp::luxAB (B) strains was measured as described by Kondo et al. (24, 26). (C and D). The ftsZp::luxAB cells were grown in LD 12:12 (125 μE/m2/s) and then released into LL, and continuous dilution of the culture was started. Cell density of the culture was monitored every hour, and a growth curve calculated from the actual cell density and the dilution rate (dilution rate = 43.3 ml of medium exchanged every hour in a total volume of 780 ml) is plotted in panel C. The average doubling time of this culture was 12.1 h. From the culture in panel C, 1 ml of cell suspension was withdrawn every 3 h and for measurement of luminescence (D) as described by Mori et al. (35).
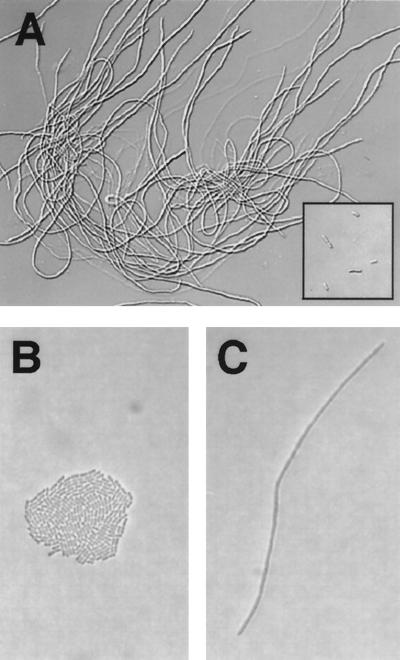
Cell division of growing cyanobacteria is stopped by overexpression of FtsZ, resulting in filamentous cells. (A) The trcp::ftsZ strain was grown for 101 h in liquid BG-11 medium supplemented with 0.5 mM IPTG. The insert in the bottom right corner of panel A shows trcp::null cells as a control under the same conditions. (B and C) The trcp::ftsZ and trcp::null cells were also grown on solid (1.5% agar) BG-11 medium supplemented with 1 mM IPTG for 48 h. A colony of trcp::null cells (B) and a filamentous trcp::ftsZ cell (C) are shown. Presumably both the colony in (panel B) and the filament in (panel C) were derived from a single initial cell.
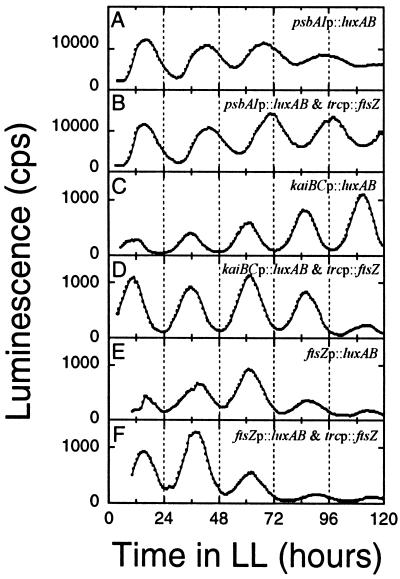
Luminescence rhythms in dividing and nondividing cyanobacteria in liquid cultures. In vivo luminescence was monitored as described by Kondo et al. (24) in the following reporter strains: (A) psbAIp::luxAB; (B) FtsZ overexpression in the psbAIp::luxAB strain; (C) kaiBCp::luxAB; (D) FtsZ overexpression in the kaiBCp::luxAB strain; (E) ftsZp::luxAB; (B) FtsZ overexpression in the ftsZp::luxAB strain. FtsZ protein was overexpressed continuously with 0.5 mM IPTG in panels B, D, and F, and filamentous morphology in those cultures was confirmed microscopically.
References
-
- Altschul S F, Gish W, Miller W, Myers E W, Lipman D J. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Andersson C R, Tsinoremas N F, Shelton J, Lebedeva N V, Yarrow J, Min H, Golden S S. Application of bioluminescence to the study of circadian rhythms in cyanobacteria. Methods Enzymol. 2000;305:527–542. - PubMed
-
- Barkai N, Leibler S. Biological rhythms: circadian clocks limited by noise. Nature. 2000;403:267–268. - PubMed
-
- Bi E F, Lutkenhaus J. FtsZ ring structure associated with division in Escherichia coli. Nature. 1991;354:161–164. - PubMed
-
- Bramhill D. Bacterial cell division. Annu Rev Cell Dev Biol. 1997;13:395–424. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
