

Vipp1 deletion mutant of Synechocystis: a connection between bacterial phage shock and thylakoid biogenesis?
- PMID: 11274448
- PMCID: PMC31210
- DOI: 10.1073/pnas.061501198
Vipp1 deletion mutant of Synechocystis: a connection between bacterial phage shock and thylakoid biogenesis?
Abstract
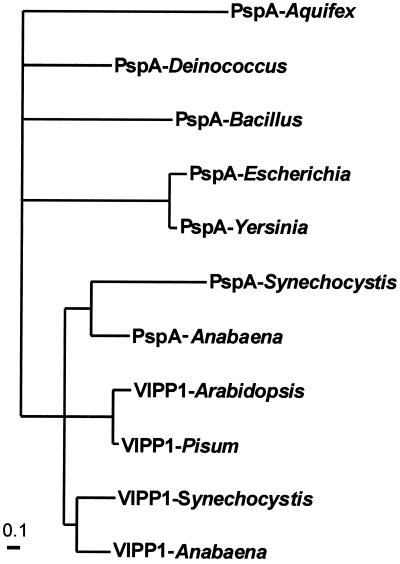
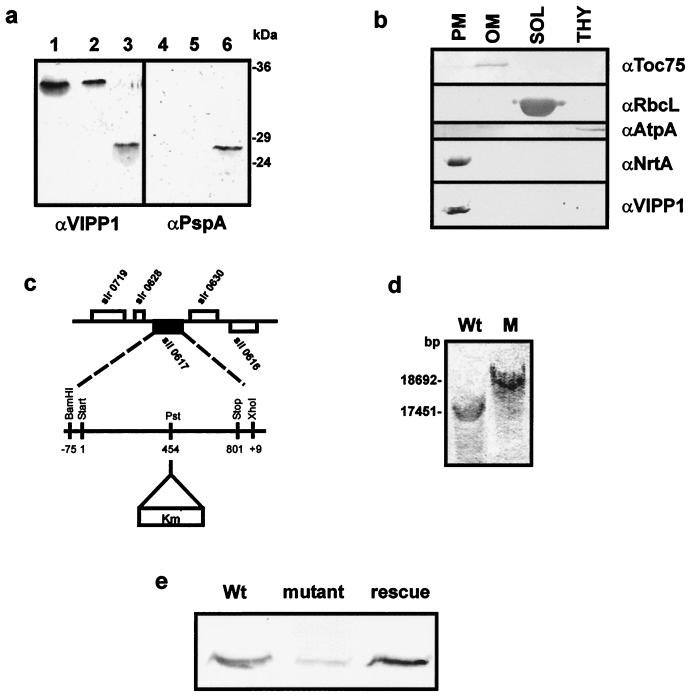
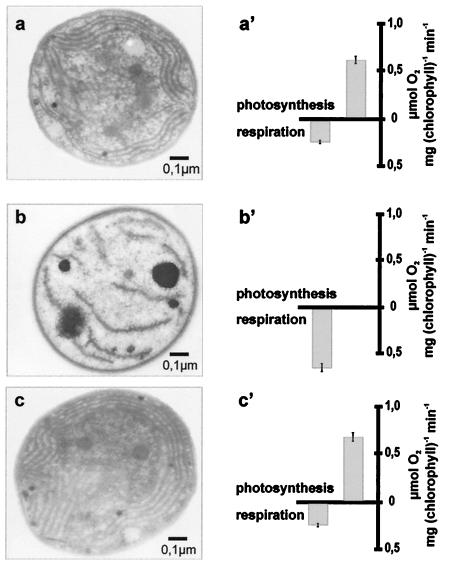
Plant chloroplasts originated from an endosymbiotic event by which an ancestor of contemporary cyanobacteria was engulfed by an early eukaryotic cell and then transformed into an organelle. Oxygenic photosynthesis is the specific feature of cyanobacteria and chloroplasts, and the photosynthetic machinery resides in an internal membrane system, the thylakoids. The origin and genesis of thylakoid membranes, which are essential for oxygenic photosynthesis, are still an enigma. Vipp1 (vesicle-inducing protein in plastids 1) is a protein located in both the inner envelope and the thylakoids of Pisum sativum and Arabidopsis thaliana. In Arabidopsis disruption of the VIPP1 gene severely affects the plant's ability to form properly structured thylakoids and as a consequence to carry out photosynthesis. In contrast, Vipp1 in Synechocystis appears to be located exclusively in the plasma membrane. Yet, as in higher plants, disruption of the VIPP1 gene locus leads to the complete loss of thylakoid formation. So far VIPP1 genes are found only in organisms carrying out oxygenic photosynthesis. They share sequence homology with a subunit encoded by the bacterial phage shock operon (PspA) but differ from PspA by a C-terminal extension of about 30 amino acids. In two cyanobacteria, Synechocystis and Anabaena, both a VIPP1 and a pspA gene are present, and phylogenetic analysis indicates that VIPP1 originated from a gene duplication of the latter and thereafter acquired its new function. It also appears that the C-terminal extension that discriminates VIPP1 proteins from PspA is important for its function in thylakoid formation.
Figures

Comment in
-
Discovery of a protein required for photosynthetic membrane assembly.Proc Natl Acad Sci U S A. 2001 Mar 27;98(7):3633-5. doi: 10.1073/pnas.071056598. Proc Natl Acad Sci U S A. 2001. PMID: 11274378 Free PMC article. No abstract available.
References
-
- Margulis L. Origin of Eukaryotic Cells. New Haven, CT: Yale Univ. Press; 1970.
-
- Kaneko T, Sato S, Kotani H, Tanaka A, Asamizu E, Nakamura Y, Miyajima N, Hirosawa M, Sugiura M, Sasamoto S, et al. DNA Res. 1996;30:109–136. - PubMed
-
- Carde J P, Joyard J, Douce R. Biol Cell. 1982;44:315–324.
-
- Douce R, Joyard R. In: Chloroplast Biogenesis. Baker N R, Barber J, editors. Amsterdam: Elsevier Science; 1984. pp. 71–132.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
